Соматическая и висцеральная мышечная система, ее фило-онтогенез. Подкожные мышцы. Скелетная мускулатура. Строение мышцы, как органа. Классификация мышц. Вспомогательные приспособления мышц.
Миология (Myologiа) - это раздел анатомии домашних животных, изучающий строение мышечной системы. Мышечная ткань, составляющая основу этой системы, осуществляет все двигательные процессы в организме животных. Благодаря ей тело фиксируется в определенном положении и перемещается в пространстве, осуществляются дыхательные движения грудной клетки и диафрагмы, движение глаз, глотание, двигательные функции внутренних органов, включая работу сердца.
Мышечная ткань обладает специальными сократительными органеллами - миофибриллами . Миофибриллы, состоящие из тонких белковых нитей (миофиламентов), могут быть неисчерченными или исчерченными (поперечно-полосатыми). Соответственно различают неисчерченную и исчерченную мышечную ткань.
1) Неисчерченная мышечная ткань состоит из клеток (гладких миоцитов) веретеновидной формы. Эти клетки образуют мышечные слои в стенках кровеносных и лимфатических сосудов, в стенках внутренних органов (желудок, кишечник, мочевыводящие пути, матка и т.д.). Длина клеток колеблется от 20 мкм (в стенке кровеносного сосуда) до 500 мкм (в стенке матки стельной коровы), диаметр от 2 до 20 мкм. В функциональном отношении неисчерченная мышечная ткань имеет ряд особенностей: она обладает большой силой (например, в кишечнике постоянно передвигаются значительные массы пищи), обладает слабой утомляемостью, медленным сокращением и ритмичностью движений (в стенке кишечника неисчерченная мышечная ткань сокращается 12 раз в минуту, а в селезенке - только 1 раз).
2) Исчерченная мышечная ткань характеризуется наличием исчерченных миофибрилл, имеет 2 разновидности.
А) Исчерченная сердечная мышечная ткань состоит из удлиненных клеток (кардиомиоцитов ) квадратной формы. Их концы, соединяясь друг с другом в цепочки, формируют так называемые функциональные мышечные " волокна" толщиной 10-20 мкм. Тесно связываясь между собой, функциональные мышечные " волокна" образуют мышечную оболочку сердца ( миокард ), постоянные и ритмичные сокращения которого приводят в движение кровь.
Б) Исчерченная скелетная мышечная ткань, в отличие от сердечной, состоит не из клеток, а из многоядерных мышечных образований (миосимпластов) цилиндрической формы. Длина миосимпластов колеблется от нескольких миллиметров до 13-15 см, диаметр от 10 до 150 мкм. Количество ядер в них может достигать нескольких десятков тысяч. Миосимпласты (их еще называют " мышечными волокнами") образуют скелетные мышцы и входят в состав некоторых органов (язык, глотка, гортань, пищевод и др.). В функциональном отношении скелетная мышечная ткань легко возбудима и сокращается быстрее, чем неисчерченная (например, в обычных условиях скелетная мышца сокращается в течение 0,1 с, а неисчерченная - в течение нескольких секунд). Но, в отличие от гладких (неисчерченных) мышц внутренних органов, скелетные мышцы быстрее утомляются.
Мышечную систему в зависимости от особенностей строения, характера двигательной функции и иннервации делят на соматическую и висцеральную.
Соматическая мышечная система составляет 40% от массы тела и построена из миосимпластов. Она произвольная и иннервируется соматической нервной системой. Соматические мышцы сокращаются быстро, энергично, но кратковременно и быстро утомляются. Такой тип сокращения называется тетаническим и он характерен для соматической мускулатуры. К ней относятся:
1) подкожные мышцы, которые не имеют связи со скелетом и прикрепляются к коже; их сокращения вызывают подергивания кожи и позволяют ей собираться в мелкие складки;
2) скелетная мускулатура, которая закрепляется на скелете;
3) диафрагма – куполообразная мышца, отделяющая грудную полость от брюшной;
4) мышцы языка, глотки, гортани, ушной раковины, глазного яблока, среднего уха, пищевода и наружных органов размножения.
Висцеральная мышечная система составляет 8% от массы тела и построена из гладких миоцитов. Она непроизвольная и иннервируется вегетативной нервной системой. Гладкие мышцы сокращаются медленно, длительно и не требуют большого количества энергии. Такой тип сокращения называют тоническим и он характерен для висцеральной мускулатуры, которая образует мышечные пучки, слои и оболочки внутренних органов.
Фило-онтогенез мышечной системы
В филогенезе хордовых мышечная система последовательно проходит ряд стадий.
У ланцетника она представлена парной продольной мышцей (правой и левой), которая идет вдоль тела и разделяется соединительнотканными перегородками (миосептами) на короткие прямые мышечные пучки (миомеры). Такое (сегментарное) деление единого мышечного пласта называется метамерией.
С увеличением подвижности, обособлением головы и развитием конечности (в виде плавников) у рыб происходит разделение продольной мышцы горизонтальной септой на дорсальную и вентральную мышцы, а так же
Обособление мускулатуры головы, туловища, хвоста и плавников.
С выходом на сушу и увеличением разнообразия движений у амфибий и рептилий происходит разделение дорсальной мышцы, так же как и вентральной, на два тяжа: латеральный (поперечно-реберная мышца) и медиальный (поперечно-остистая м.). Кроме этого, у рептилий из латерального тяжа впервые появляются подкожные мышцы, которые прикрепляются к коже.
У более высокоорганизованных животных ( птиц и млекопитающих ) происходит дальнейшая дифференциация мышечной системы: латеральный и медиальный тяжи, каждый из них, разделяются на два слоя (поверхностный и глубокий). Кроме этого, у млекопитающих впервые появляется диафрагма.
Филогенез мышечной системы.
| Хордовые | Мышечная система | |||||||
| Ланцетник | Продольная мышца | |||||||
| Рыбы | Дорсальная | Вентральная | ||||||
| Амфибии, рептилии | Латеральная | Медиальная | Латеральная | Медиальная | ||||
| Птицы, млекопитающие | Повер | Глуб. | П | Г | П | Г | П | Г |
В онтогенезе мышечная система в основном развивается из миотомов мезодермы, исключение составляют некоторые мышцы головы и шеи, кторые образуются из мезенхимы (трапециевидная, плечеголовная).
В начале формируется мышечный продольный тяж, который сразу же дифференцируется на дорсальный и вентральный пласты; далее каждый из них разделяется на латеральный и медиальный пласты, которые, в свою очередь, дифференцируются на поверхностный и глубокий слои, последние дают начало определенным группам мышц. Например, из поверхностного слоя латерального пласта развиваются подвздошно-реберные мышцы, а из глубокого слоя латерального пласта – длиннейшие мышцы спины, шеи, головы.
Подкожные мышцы – musculi cutanei
Подкожные мышцы прикрепляются к кожному покрову, фасциям и не имеют связи со скелетом. Их сокращения вызывают поддергивания кожи и позволяют ей собираться в мелкие складки. К этим мышцам относятся:
1) Подкожная мышца шеи – m. Cutaneus colli (особенно сильно развита у собак). Она идет вдоль шеи, ближе к ее вентральной поверхности и переходит на лицевую поверхность к мышцам рта и нижней губы.
2) Подкожная мышца лопатки и плеча (лопаточно-плечевая) – m. Cutaneus omobrachialis. Она покрывает область лопатки и частично плечо. Хорошо выражена у лошади и крупного рогатого скота.
3) Подкожная мышца туловища – m. Cutaneus trunci. Она расположена по бокам от грудной и брюшной стенок и каудально отдает пучки в коленную складку.
4) У самок в области молочных желез располагаются краниальные и каудальные мышцы молочной железы (mm. Supramammilaris cranialis et caudalis), которые придают складчатость коже и помогают выведению молока. Сильно развиты у хищных животных.
У самцов в этой области имеются краниальные и каудальные препуциальные мышцы (mm.preputialis cranialis et caudalis), которые обеспечивают складчатость препуция и выполняют роль его сфинктера.
Скелетная мускулатура
Скелетная мускулатура представляет собой активную часть опорно-двигательного аппарата. Она состоит из скелетных мышц и их вспомогательных приспособлений, к которым относятся фасции, синовиальные сумки, синовиальные влагалища сухожилий, блоки, сезамовидные кости.
В теле животного насчитывается около 500 скелетных мышц. Большинство из них парные и располагаются симметрично по обеим сторонам тела животного. Их суммарная масса составляет у лошади 38-42% от массы тела, у крупного рогатого скота 42-47%, у свиней 30-35% от массы тела.
Мышцы в теле животного располагаются не беспорядочно, а закономерно в зависимости от действия силы тяжести животного и выполняемой работы. Они оказывают свое действие на те части скелета, которые соединены подвижно, т.е. мышцы действуют на суставы, синдесмозы.
Основными местами прикрепления мышц являются кости, но иногда они прикрепляются к хрящам, связкам, фасциям, коже. Они покрывают скелет так, что кости лишь в некоторых местах лежат непосредственно под кожей. Закрепляясь на скелете, как на системе рычагов, мышцы при своем сокращении вызывают различные движения тела, фиксируют скелет в определенном положении и придают форму телу животного
Основные функции скелетных мышц:
1) Основная функция мышц - динамическая. Сокращаясь, мышца укорачивается на 20-50% своей длины и тем самым меняет положение связанных с ней костей. Производится работа, результатом которой является движение.
2) Другая функция мышц – статическая. Проявляется она в фиксации тела в определенном положении, в сохранении формы тела и его частей. Одна из проявлений этой функции – способность спать стоя (лошадь).
3) Участие в обмене веществ и энергии. Скелетные мышцы являются «источниками тепла», так как при их сокращении около 70% энергии превращается в тепло и только 30% энергии обеспечивает движение. В скелетных мышцах удерживается около 70% воды организма, поэтому их еще называют «источниками воды». Кроме этого, между мышечными пучками и внутри их может накапливаться жировая ткань (особенно при откорме у свиней).
4) Одновременно, при своей работе скелетные мышцы помогают работе сердца, проталкивая венозную кровь по сосудам. В экспериментах удалось выяснить, что скелетные мышцы действуют подобно насосу, обеспечивая движение крови по венозному руслу. Поэтому скелетные мышцы еще называют «периферическими мышечными сердцами».
Строение мышцы с точки зрения биохимика
Скелетная мышца состоит из органических и неорганических соединений. К неорганическим соединениям относятся вода и минеральные соли (соли кальция, фосфора, магния). Органическое вещество в основном представлено белками, углеводами (гликоген), липидами (фосфатиды, холестерин).
Таблица 2. Химический состав скелетной мышцы
Химический состав скелетных мышц подвержен значительным возрастным и в меньшей степени видовым, породным и половым отличиям, что прежде всего связано с неодинаковым содержанием в них воды (с возрастом % воды уменьшается).
Отправить свою хорошую работу в базу знаний просто. Используйте форму, расположенную ниже
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
Размещено на http://www.allbest.ru/
Введение
Миология (Myologiа) - это раздел анатомии домашних животных, изучающий строение мышечной системы. Мышечная ткань, составляющая основу этой системы, осуществляет все двигательные процессы в организме животных. Благодаря ей тело фиксируется в определенном положении и перемещается в пространстве, осуществляются дыхательные движения грудной клетки и диафрагмы, движение глаз, глотание, двигательные функции внутренних органов, включая работу сердца.
Мышечная ткань обладает специальными сократительными органеллами - миофибриллами. Миофибриллы, состоящие из тонких белковых нитей (миофиламентов), могут быть неисчерченными или исчерченными (поперечно-полосатыми). Соответственно различают неисчерченную и исчерченную мышечную ткань.
1) Неисчерченная мышечная ткань состоит из клеток (гладких миоцитов) веретеновидной формы. Эти клетки образуют мышечные слои в стенках кровеносных и лимфатических сосудов, в стенках внутренних органов (желудок, кишечник, мочевыводящие пути, матка и т.д.). Длина клеток колеблется от 20 мкм (в стенке кровеносного сосуда) до 500 мкм (в стенке матки стельной коровы), диаметр от 2 до 20 мкм. В функциональном отношении неисчерченная мышечная ткань имеет ряд особенностей: она обладает большой силой (например, в кишечнике постоянно передвигаются значительные массы пищи), обладает слабой утомляемостью, медленным сокращением и ритмичностью движений (в стенке кишечника неисчерченная мышечная ткань сокращается 12 раз в минуту, а в селезенке - только 1 раз).
2) Исчерченная мышечная ткань характеризуется наличием исчерченных миофибрилл, имеет 2 разновидности.
А) Исчерченная сердечная мышечная ткань состоит из удлиненных клеток (кардиомиоцитов) квадратной формы. Их концы, соединяясь друг с другом в цепочки, формируют так называемые функциональные мышечные "волокна" толщиной 10-20 мкм. Тесно связываясь между собой, функциональные мышечные "волокна" образуют мышечную оболочку сердца (миокард), постоянные и ритмичные сокращения которого приводят в движение кровь.
Б) Исчерченная скелетная мышечная ткань, в отличие от сердечной, состоит не из клеток, а из многоядерных мышечных образований (миосимпластов) цилиндрической формы. Длина миосимпластов колеблется от нескольких миллиметров до 13-15 см, диаметр от 10 до 150 мкм. Количество ядер в них может достигать нескольких десятков тысяч. Миосимпласты (их еще называют "мышечными волокнами") образуют скелетные мышцы и входят в состав некоторых органов (язык, глотка, гортань, пищевод и др.). В функциональном отношении скелетная мышечная ткань легко возбудима и сокращается быстрее, чем неисчерченная (например, в обычных условиях скелетная мышца сокращается в течение 0,1 с, а неисчерченная - в течение нескольких секунд). Но, в отличие от гладких (неисчерченных) мышц внутренних органов, скелетные мышцы быстрее утомляются.
Мышечную систему в зависимости от особенностей строения, характера двигательной функции и иннервации делят на соматическую и висцеральную.
Соматическая мышечная система составляет 40% от массы тела и построена из миосимпластов. Она произвольная и иннервируется соматической нервной системой. Соматические мышцы сокращаются быстро, энергично, но кратковременно и быстро утомляются. Такой тип сокращения называется тетаническим и он характерен для соматической мускулатуры. К ней относятся:
1) подкожные мышцы, которые не имеют связи со скелетом и прикрепляются к коже; их сокращения вызывают подергивания кожи и позволяют ей собираться в мелкие складки;
2) скелетная мускулатура, которая закрепляется на скелете;
3) диафрагма - куполообразная мышца, отделяющая грудную полость от брюшной;
4) мышцы языка, глотки, гортани, ушной раковины, глазного яблока, среднего уха, пищевода и наружных органов размножения.
Висцеральная мышечная система составляет 8% от массы тела и построена из гладких миоцитов. Она непроизвольная и иннервируется вегетативной нервной системой. Гладкие мышцы сокращаются медленно, длительно и не требуют большого количества энергии. Такой тип сокращения называют тоническим и он характерен для висцеральной мускулатуры, которая образует мышечные пучки, слои и оболочки внутренних органов.
1. Фило-онтогенез мышечной системы
В филогенезе хордовых мышечная система последовательно проходит ряд стадий.
У ланцетника она представлена парной продольной мышцей (правой и левой), которая идет вдоль тела и разделяется соединительнотканными перегородками (миосептами) на короткие прямые мышечные пучки (миомеры). Такое (сегментарное) деление единого мышечного пласта называется метамерией.
С увеличением подвижности, обособлением головы и развитием конечности (в виде плавников) у рыб происходит разделение продольной мышцы горизонтальной септой на дорсальную и вентральную мышцы.
Обособление мускулатуры голов ы, туловища, хвоста и плавников
С выходом на сушу и увеличением разнообразия движений у амфибий и рептилий происходит разделение дорсальной мышцы, так же как и вентральной, на два тяжа: латеральный (поперечно-реберная мышца) и медиальный (поперечно-остистая м.). Кроме этого, у рептилий из латерального тяжа впервые появляются подкожные мышцы, которые прикрепляются к коже.
У более высокоорганизованных животных (птиц и млекопитающих) происходит дальнейшая дифференциация мышечной системы: латеральный и медиальный тяжи, каждый из них, разделяются на два слоя (поверхностный и глубокий). Кроме этого, у млекопитающих впервые появляется диафрагма.
Филогенез мышечной системы
В онтогенезе мышечная система в основном развивается из миотомов мезодермы, исключение составляют некоторые мышцы головы и шеи, кторые образуются из мезенхимы (трапециевидная, плечеголовная).
В начале формируется мышечный продольный тяж, который сразу же дифференцируется на дорсальный и вентральный пласты; далее каждый из них разделяется на латеральный и медиальный пласты, которые, в свою очередь, дифференцируются на поверхностный и глубокий слои, последние дают начало определенным группам мышц. Например, из поверхностного слоя латерального пласта развиваются подвздошно-реберные мышцы, а из глубокого слоя латерального пласта - длиннейшие мышцы спины, шеи, головы.
2. Подкожные мышцы - musculi cutanei
Подкожные мышцы прикрепляются к кожному покрову, фасциям и не имеют связи со скелетом. Их сокращения вызывают поддергивания кожи и позволяют ей собираться в мелкие складки. К этим мышцам относятся:
1) Подкожная мышца шеи - m. Cutaneus colli (особенно сильно развита у собак). Она идет вдоль шеи, ближе к ее вентральной поверхности и переходит на лицевую поверхность к мышцам рта и нижней губы.
2) Подкожная мышца лопатки и плеча (лопаточно-плечевая) - m. Cutaneus omobrachialis. Она покрывает область лопатки и частично плечо. Хорошо выражена у лошади и крупного рогатого скота.
3) Подкожная мышца туловища - m. Cutaneus trunci. Она расположена по бокам от грудной и брюшной стенок и каудально отдает пучки в коленную складку.
4) У самок в области молочных желез располагаются краниальные и каудальные мышцы молочной железы (mm. Supramammilaris cranialis et caudalis), которые придают складчатость коже и помогают выведению молока. Сильно развиты у хищных животных.
У самцов в этой области имеются краниальные и каудальные препуциальные мышцы (mm.preputialis cranialis et caudalis), которые обеспечивают складчатость препуция и выполняют роль его сфинктера.
3. Скелетная мускулатура
Скелетная мускулатура представляет собой активную часть опорно-двигательного аппарата. Она состоит из скелетных мышц и их вспомогательных приспособлений, к которым относятся фасции, синовиальные сумки, синовиальные влагалища сухожилий, блоки, сезамовидные кости.
В теле животного насчитывается около 500 скелетных мышц. Большинство из них парные и располагаются симметрично по обеим сторонам тела животного. Их суммарная масса составляет у лошади 38-42% от массы тела, у крупного рогатого скота 42-47%, у свиней 30-35% от массы тела.
Мышцы в теле животного располагаются не беспорядочно, а закономерно в зависимости от действия силы тяжести животного и выполняемой работы. Они оказывают свое действие на те части скелета, которые соединены подвижно, т.е. мышцы действуют на суставы, синдесмозы.
Основными местами прикрепления мышц являются кости, но иногда они прикрепляются к хрящам, связкам, фасциям, коже. Они покрывают скелет так, что кости лишь в некоторых местах лежат непосредственно под кожей. Закрепляясь на скелете, как на системе рычагов, мышцы при своем сокращении вызывают различные движения тела, фиксируют скелет в определенном положении и придают форму телу животного
Основные функции скелетных мышц:
1) Основная функция мышц - динамическая. Сокращаясь, мышца укорачивается на 20-50% своей длины и тем самым меняет положение связанных с ней костей. Производится работа, результатом которой является движение.
2) Другая функция мышц - статическая. Проявляется она в фиксации тела в определенном положении, в сохранении формы тела и его частей. Одна из проявлений этой функции - способность спать стоя (лошадь).
3) Участие в обмене веществ и энергии. Скелетные мышцы являются «источниками тепла», так как при их сокращении около 70% энергии превращается в тепло и только 30% энергии обеспечивает движение. В скелетных мышцах удерживается около 70% воды организма, поэтому их еще называют «источниками воды». Кроме этого, между мышечными пучками и внутри их может накапливаться жировая ткань (особенно при откорме у свиней).
4) Одновременно, при своей работе скелетные мышцы помогают работе сердца, проталкивая венозную кровь по сосудам. В экспериментах удалось выяснить, что скелетные мышцы действуют подобно насосу, обеспечивая движение крови по венозному руслу. Поэтому скелетные мышцы еще называют «периферическими мышечными сердцами».
4. Строение мышцы с точки зрения биохимика
Скелетная мышца состоит из органических и неорганических соединений. К неорганическим соединениям относятся вода и минеральные соли (соли кальция, фосфора, магния). Органическое вещество в основном представлено белками, углеводами (гликоген), липидами (фосфатиды, холестерин).
Химический состав скелетной мышцы
Химический состав скелетных мышц подвержен значительным возрастным и в меньшей степени видовым, породным и половым отличиям, что прежде всего связано с неодинаковым содержанием в них воды (с возрастом % воды уменьшается).
5. Строение мышцы с точки зрения анатома
Скелетная мышца (Musculus skeleti) - это активный орган аппарата движения, форма и особенности строения которого обусловлены выполняемой функцией и местоположением на скелете. В мышце различают активно сокращающуюся часть - мышечное брюшко и пассивную часть, при помощи которой она прикрепляется к костям, - сухожилие.
1) Мышечное брюшко (venter) состоит из паренхимы и стромы. Паренхима представлена исчерченной мышечной тканью, структурной единицей которой является миосимпласт. Миосимпласты объединяются при помощи рыхлой соединительной ткани, которая называется эндомизий, в пучки 1 порядка. Пучки 1 порядка объединяются в пучки 1,2,3 пордка и между ними формируются соединительнотканные перегородки (перимизий), по которым внутрь в мышцу проникают сосуды и нервы. Снаружи мышечное брюшко покрыто соединительнотканной оболочкой (эпимизием). Эндо-, пери- и эпимизий образуют строму мышечного брюшка и защищают мышцу от чрезмерного утолщения или растяжения. Соединительнотканные элементы, имеющиеся между мышечными волокнами, по концам мышечного брюшка, переходят в сухожилия.
2) Сухожилие (tendo) построено по тому же принципу, что и мышечное брюшко, с той лишь разницей, что вместо мышечных волокон его пучки содержат коллагеновые волокна. Прослойки соединительной ткани внутри носят названия эндо- и пери теноний, а снаружи плотная соединительная ткань образует оболочку (эпитеноний), которая является продолжением эпимизия. Сухожилие имеет блестящий светло-золотистый цвет, резко отличающийся от красно-бурого цвета брюшка мышцы. В большинстве случаев сухожилие находится по обоим концам мышцы и прикрепляется к костям. Хотя сухожилие значительно тоньше мышечного брюшка, прочность его велика, оно способно выдерживать большую нагрузку и практически нерастежимо. Исследования показали, что для разрыва ахиллового сухожилия у животного требуется сила от 900 кг на один кубический см.
3) Сосуды и нервы входят в мышцу с ее внутренней стороны.
Артерии ветвятся до капилляров, которые в пучках мышечных волокон образуют густую сеть. К каждому мышечному волокну прилежит не менее одного кровеносного капилляра. В каждую мышцу кровь поступает по артериям, а оттекает по венам и лимфатическим сосудам.
Нервы, разветвляясь в мышце, образуют нервно-мышечный комплекс - мион, который состоит из 1 нервного волокна и нескольких мышечных волокон. Так, например, в трехглавой мышце голени мион состоит из 1нервного волокна и 227 мышечных волокон, а в латеральной мышце глаза - из 1 нервного волокна и 19 мышечных волокон.
Рост мышц в длину происходит в так называемых «зонах роста», которые располагаются в местах перехода мышечного брюшка в сухожилие и содержат большое количество ядер, а увеличение мышц в толщину происходит благодаря функциональной нагрузке, которую выполняет данная мышца.
6. Классификация мышц
Каждая мышца является самостоятельным органом и имеет определенную форму, величину, строение, функцию, происхождение и положение в организме. В зависимости от этого все скелетные мышцы подразделяются на следующие группы.
I. По форме различают мышцы длинные, короткие, плоские и т.д.
1) Длинные мышцы соответствуют длинным рычагам движения и поэтому встречаются главным образом на конечностях. Имеют веретенообразную форму, средняя часть называется брюшком, конец, соответствующий началу мышцы, - головкой, противоположный конец - хвостом. Сухожилие длинных мышц имеет форму ленты. Некоторые длинные мышцы начинаются несколькими головками (многоглавые) на различных костях, что усиливает их опору. Встречаются мышцы двуглавые (двуглавая м. плеча), трехглавые (трехглавая м. голени) и четырехглавые (четырехглавая м бедра).
2) Короткие мышцы находятся на тех участках тела, где размах движений невелик (между отдельными позвонками (многораздельные м.), между позвонками и ребрами (подниматели ребер) и т.д.).
3) Плоские (широкие) мышцы располагаются преимущественно на туловище и поясах конечностей. Они имеют расширенное сухожилие, называемое апоневрозом. Плоские мышцы обладают не только двигательной функцией, но также опорной и защитной (например, мышцы брюшной стенки защищают и способствуют удержанию внутренних органов).
4) Встречаются также и другие формы мышц: квадратная, круговая, дельтовидная, зубчатая, трапецевидная, веретеновидная и др.
II. По анатомическому строению мышцы делятся в зависимости от количества внутримышечных сухожильных прослоек и направления мышечных прослоек:
1) Одноперистые. Для них характерно отсутствие сухожильных прослоек и мышечные волокна присоединяются к сухожилию одной стороны (наружная косая брюшная м.).
2) Двуперистые. Для них характерно наличие одной сухожильной прослойки и мышечные волокна присоединяются к сухожилию с двух сторон (трапецевидная м.).
3) Многоперистые. Для них характерно наличие двух и более сухожильных прослоек, в результате этого мышечные пучки сложно переплетаются и к сухожилию подходят с нескольких сторон (жевательная м., дельтовидная мышца).
III. По гистоструктуре все мышцы делятся на 3 типа в зависимости от соотношения исчерченной мышечной ткани к соединительной:
1) Динамический тип. Для динамических мышц, обеспечивающих активную и разностороннюю работу, характерно значительное преобладание исчерченной мышечной ткани над соединительной (четырехглавая м. бедра).
2) Статический тип. В отличие от динамических, статические мыщцы совсем не имеют мышечных волокон. Они выполняют большую статическую работу при стоянии и опоре конечности о почву во время движения, закрепляя суставы в определенном положении (третья межкостная м. коровы и лошади)
3) Статодинамический тип. Для этого типа характерно уменьшение отношения исчерченной мышечной ткани к соединительно-тканным элементам (двуглавая м. плеча лошади). Статодинамические мышцы, как правило, имеют перистое строение.
IV. По действию на суставы мышцы делятся на одно-, дву- и многосуставные.
1) Односуставные действуют только на один сустав (предостная м., заостная м. действуют на плечевой сустав).
2) Двусуставные, действуют на два сустава (напрягатель широкой фасции бедра действует на тазобедренный и коленный суставы).
3) Многосуставные (двуглавая м. бедра, полусухожильная м., полуперепончатая м. действуют на 3 сустава (тазобедренный, коленный, скакательный).
Кроме того, необходимо подчеркнуть, что мышцы действуют раздельно или группой. Одинаково действующие мышцы называются синергистами, а действующие противоположным образом - антагонистами.
V. По функции мышцы делятся на:
1. Флексоры, или сгибатели, которые при сокращении сближают концы костей 2. Экстензоры, или разгибатели, которые проходят через вершину угла сустава и при сокращении раскрывают его.
3. Абдукторы, или отводящие мышцы, лежат на латеральной стороне сустава и отводят его от сагиттальной плоскости в сторону.
4. Аддукторы, или приводящие мышцы, лежат на медиальной поверхности сустава и при сокращении приводят его к сагиттальной плоскости.
5. Ротаторы, или вращатели, обеспечивающие вращение конечности наружу (супинаторы) или во внутрь (пронаторы).
6. Сфинктеры, или запиратели, которые располагаются вокруг естественных отверстий и при сокращении закрывают их. Для них, как правило, характерно круговое направление мышечных волокон (например, круговая мышца рта).
7. Констрикторы, или суживатели, которые также относятся к типу круглых мышц, но имеют иную форму (например, констрикторы глотки, гортани).
8. Дилататоры, или расширители, при сокращении открывают естественные отверстия.
9. Леваторы, или подниматели при сокращение поднимают, например ребра.
10. Депрессоры, или опускатели.
11. Тензоры, или напрягатели, своей работой напрягают фасции, не позволяя им собираться в складки.
12. Фиксаторы, укрепляют сустав на стороне расположения соответствующих мышц.
VI. По происхождению все скелетные мышцы делятся на соматические и висцеральные.
1) Соматические мышцы развиваются из сомитов мезодермы (жевательная м., височная м., м. позвоночного столба).
2) Висцеральные являются производными мышц жаберного аппарата. К висцеральной мускулатуре относятся мышцы головы (мимические, жевательные) и некоторые мышцы шеи.
мышечный система человек животное
7. Вспомогательные приспособления мышц
Мышцы сокращаясь, выполняют свою функцию при участии и при помощи анатомических образований, которые следуют рассматривать как вспомогательные приспособления мышц. Они улучшают работу мышц. К ним относятся фасции, сумки, синовиальные влагалища сухожилий, блоки и сезамовидные кости.
Фасция (лат.fascia- обертка)
Фасции - это тонкие, прочные, соединительнотканные оболочки, которые образуют вокруг мышц своеобразные футляры. Они в основном выполняют опорную и амортизационную функции. Фасции отграничивают мышцы друг от друга, создают опору для мышечного брюшка при его сокращении и устраняют трение мышц друг от друга. Фасции еще называют мягким скелетом (считают остатком перепончатого скелета предков позвоночных). Они богаты нервными окончаниями (рецепторами) и сосудами и поэтому играют существенную роль в восстановительных (регенерационных) процессах. Так, например, если при удалении пораженного мениска в коленном суставе на его место приживить лоскут фасции, не потерявшей связи с сосудами и нервами, то при определенной тренировке через некоторое время на её месте сформируется «орган» наподобие мениска и работа сустава в целом восстанавливается. Поэтому фасции широко используются в реконструктивной хирургии при аутопластике хрящевой и костной тканей. Фасции бывают поверхностные, глубокие и специальные фасции.
Поверхностные, или подкожные, фасции отделяют кожный покров от скелетной мускулатуры и образуют своеобразные футляры для всех областей тела животного. К ним прикрепляются подкожные мышцы.
1) Поверхностная ф головы (f.superficialis capitis), в ней заключены мышцы головы.
2) Шейная ф. (f.cervicalis) лежит вентрально в области шеи и прикрывает трахею.
3) Грудопоясничная ф.(f.thoracolubalis) лежит дорсально на туловище и закрепляется на остистых отростках грудных и поясничных позвонков и маклоке.
4) Грудобрюшная ф. (f.thoracoabdominalis) лежит латерально по бокам от грудной и брюшной полости и закрепляется вентрально по белой линии живота (linea alba).
5) Поверхностная ф. грудной конечности (f.superficialis membri thoracici) является продолжением грудобрюшной фасции. Она значительно утолщена в области запястья и формирует фиброзные влагалища для сухожилий мышц, которые здесь проходят.
6) Поверхностная ф. тазовой конечности (f.superficialis membri pelvini) является продолжением грудопоясничной и значительно утолщена в области заплюсны.
Глубокие, или собственные, фасции прикрепляются к костям и удерживают мышцы в определенном положении, не давая им смещаться. Они образуют футляры для отдельных мышц, групп мышц (синергистов) и органов.
1) В области головы поверхностная фасция делится на следующие глубокие: лобную (покрывает спинку носа), височную, околоушно-жевательную, щечную, подчелюстную, щечно-глоточную.
2) Внутригрудная (f.endothoracica) выстилает внутреннюю поверхность грудной полости.
3) Поперечно-брюшная (f.transversalis) выстилает внутреннюю поверхность брюшной полости.
4) Тазовая (f.pelvis) выстилает внутреннюю поверхность тазовой полости.
5) В области грудной конечности поверхностная фасция делится на следующие глубокие: фасции лопатки, плеча, предплечья, кисти, пальцев.
6) В области тазовой конечности поверхностная фасция делится на следующие глубокие: ягодичную (покрывает область крупа), фасции бедра, голени, стопы, пальцев
Специальные покрывают отдельные мышцы. Например, глубокая околоушно-жевательная фасция делится на две специальные: околоушная покрывает слюнную железу, а жевательная - жевательную мышцу.
Бурса (bursa - сумка)
В местах прикрепления и наибольшей подвижности сухожилий и мышц имеются бурсы. Они имеют форму плоского соединительно-тканного мешочка, внутри которого находится жидкость. Бурсы уменьшают трение и смягчают сопрокосновение мышц с другими органами (костью, кожей). Они имеют различную величину: от нескольких миллиметров до нескольких сантиметров. В зависимости от того, чем заполнены бурсы, различают синовиальные и слизистые бурсы.
1) Синовиальные бурсы (bursa synovialis) образуются за счет капсулы сустава и заполнены синовией, поэтому полость бурсы сообщается с полостью сустава. Такие бурсы расположены в основном в области локтевого и коленного сустава. Воспаление этих бурс вследствие травмы может привести к артриту (воспалению сустава) локтевого или коленного суставов, а об этом необходимо помнить в ветеринарной практике.
2) Cлизистые бурсы (bursa mucosa) образуются в уязвимых местах под связками (подсвязочные), под мышцами (подмышечные), под сухожилиями (подсухожильные) и под кожей (подкожные). Полость их заполнена слизью и они могут быть постоянными или временными (мозоли).
Синовиальное влагалище сухожилий (vagina synovialis tendinis )
Синовиальное влагалище сухожилий отличается от синовиальной сумки тем, что имеет гораздо большие размеры (длину, ширину) и двойную стенку. Оно полностью охватывает движущееся в нем сухожилие мышцы, которое заключено как бы в трубку, заполненную синовией. Вследствие этого синовиальное влагалище не только выполняет функцию бурсы, но и укрепляет положение сухожилия мышцы на значительном ее протяжении. Встречаются в области запястного, заплюсневого и пальцевых суставов.
Синовиальное влагалище ограничено листками. Висцеральный (внутренний) листок окружает со всех сторон сухожилие и срастается с ним. Париетальный (наружный) выстилает стенки фиброзного влагалища. Оба листка переходят в друг друга на концах влагалища и вдоль его сухожилия. Удвоенный листок влагалища, соединяющий внутренний и наружный листки называется брыжейкой сухожилия или мезотендинием.
Блок (trochlea)
Блоки - это определенной формы участки эпифизов трубчатых костей, через которые перекидываются мышцы. Они представляет собой костный выступ и желобок в нем, где проходит сухожилие мышц. Благодаря этому сухожилия не смещаются в сторону и увеличивается рычаг приложения силы. Какие кости имеют блоки? Плечевая, бедренная.
Сезамовидные кости (ossa sesamoidea)
Сезамовидные кости формируются в области очень сильного напряжения мышц и обнаруживаются в толще сухожилий. Они изменяют угол прикрепления мышц и тем самым улучшают условия их работы, уменьшая трение. Иногда их называют «окостеневшими участками сухожилий», но необходимо помнить, что они проходят только две стадии развития (соединительнотканную и костную).
Самая крупная сезамовидная кость организма - коленная чашечка.
Список литературы
1. Агаджанян Н.А., Власова И.Г., Ермакова Н.В., Трошин В.И. Основы физиологии человека: Учебник - М., 2009.
2. Антонова В.А. Возрастная анатомия и физиология. - М.: Высшее образование. - 192 с. 2006.
3. Воробьева Е.А. Анатомия и физиология. - М.: Медицина, 2007.
4. Липченко В.Я. Атлас нормальной анатомии человека. - М.: Медецина. 2009.
5. Обреумова Н.И., Петрухин А.С. Основы анатомии, физиологии и гигиены детей и подростков. Учебное пособие для студентов дефектологический факультете высш. пед. учеб. заведений. - М.: Издательский центр «Академия», 2008.
Размещено на Allbest.ru
...Подобные документы
Значение мышечной системы в жизнедеятельности организма человека. Строение скелетных мышц, основные группы и гладкие мышцы и их работа. Характеристика основных групп скелетных мышц. Возрастные особенности мышечной системы. Мышцы руки, кисти и голени.
презентация , добавлен 11.12.2014
Строение и функции суставов, позвоночника, скелетных мышц. Основные группы мышц и особенности их работы. Возрастные изменения костно-мышечной системы. Последствия гиподинамии, ключевые фазы и виды работоспособности человека. Проблема снятия переутомления.
реферат , добавлен 14.01.2014
Изучение особенностей строения и функций мышц - активной части двигательного аппарата человека. Характеристика мышц туловища, фасций спины (поверхностных и глубоких), груди, живота, головы (мышцы лица, жевательные мышцы). Физиологические свойства мышц.
реферат , добавлен 23.03.2010
Произвольные и непроизвольные мыщцы. Отведение и вращение внутрь – основные функции мышц. Свойства мышечной ткани: возбудимость, сократимость, растяжимость, эластичность. Функции скелетных (соматических) мышц. Особенности мышц синергистов и антагонистов.
презентация , добавлен 13.12.2010
Понятие скелетной (соматической) мускулатуры, ее структура и элементы. Содержание в мышцах сосудов и нервов, их роль и значение в нормальной деятельности мышц. Классификация мышц по форме, внутренней структуре и действию, их виды и характеристика.
контрольная работа , добавлен 09.02.2009
Исследование расположения и основных функций поверхностных и глубоких мышц груди. Описания мышечных пучков диафрагмы. Прикрепление пирамидальной, поперечной, прямой мышц живота и квадратной мышцы поясницы. Внутренние межреберные и подреберные мышцы.
презентация , добавлен 18.04.2015
Терморегуляция, строение и значение кожи. Система опоры и движения, скелет. Мышцы, их строение, функции и работа. Развитие организма человека. Размножение в органическом мире. Беременность, развитие зародыша и плода. Развитие человека после рождения.
реферат , добавлен 06.07.2010
Строение таза - расположенной в основании позвоночника части скелета человека. Поперечные размеры измерения таза. Связь формы и величины таза с его функцией. Подвздошно-поясничная, внутренняя запирательная, грушевидная мышцы. Строение женского таза.
презентация , добавлен 18.03.2015
Основные элементы и химический состав мышечной ткани. Виды белков саркоплазмы и миофибрилл, их содержание к общему количеству белков, молекулярная масса, распределение в структурных элементах мышцы. Их функции и роль организме. Строение молекулы миозина.
презентация , добавлен 14.12.2014
Опорно-двигательная функция нижней конечности. Передняя и задняя группы мышц голени. Длинный разгибатель пальцев. Трехглавая мышца голени. Мышцы тыла стопы. Икроножная, камбаловидная, подошвенная и подколенная мышцы. Задняя большеберцовая мышца.
В середине диафиза трубчатой кости имеется костная полость, распространяющаяся у взрослых животных до эпифизов. Она образовалась в результате резорбции костной ткани остеокластами во время развития кости и заполнена желтым (жировым) костным мозгом. Стенки костной полости выстланы эндостом, выполняющим, как и периост, камбиальную функцию. Им же покрыты перекладины губчатого вещества.
Кость богата сосудами, которые образуют сеть в ее надкостнице, пронизывают всю толщину компактного вещества, находясь в центре каждого остеона, и разветвляются в костном мозге. В кости кроме сосудов остеонов имеются так называемые питательные сосуды (фолькмановы), прободающие кость перпендикулярно ее длине. Вокруг них не образуется концентрических костных пластинок. Особенно много таких сосудов около эпифизов. Нервы в кость проникают от надкостницы через те же отверстия, что и сосуды. Поверхность кости, обращенная в полость сустава, покрыта гиалиновым хрящом без надхрящницы. Толщина его 0,2-6 мм и прямо пропорциональна нагрузке на сустав.
Строение коротких, сложных и плоских костей такое же, как и трубчатых, с той лишь разницей, что у них обычно нет костных полостей. Исключение составляют некоторые плоские кости головы, в которых между пластинками компактного вещества имеются обширные пространства, заполненные воздухом,- синусы или пазухи.
ФИЛОГЕНЕЗ СКЕЛЕТА
Развитие опорной системы в филогенезе многоклеточных животных шло двумя путями: образования наружного и внутреннего скелета. Наружный скелет закладывается в покровах тела. Наивысшего развития достигает у членистоногих. Внутренний скелет развивается вне связи с покровами тела, под кожей и обычно прикрыт мышцами. Его элементы встречаются и у беспозвоночных, но о развитии внутреннего скелета можно говорить со времени появления хордовых. У примитивных хордовых (оболочники, ланцетник)- спинная струна - хорда представляет собой опорную систему. У бесчерепных (ланцетник) внутренний скелет дополняется соединительнотканным футляром хорды, от которого отходят пластинки - миосепты, разделяющие мышечные пласты на отдельные сегменты (у ланцетника их около
С усложнением организации животных соединительнотканный скелет замещается хрящевым, а затем костным. Наряду со сменой тканевой основы скелета происходило и его структурное усложнение.
Филогенез стволового скелета. В филогенезе позвоночных раньше других элементов скелета появляются позвонки. У наиболее примитивных позвоночных- круглоротых (миноги) зачатки позвонков развиваются в виде хрящевых дуг, метамерно расположенных над хордой, выполняющей роль основного опорного стержня тела (рис. 41-А -Д). Хрящевые дуги по своему строению однотипны, так как движения круглоротых однообразны и муску-
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
латура недифференцирована на отдельные мышцы, в результате чего осевой скелет нечетко разделяется на головной, туловищный и хвостовой отделы.
Рис. 41. Схема последовательных стадий образования позвонка.
Схема позвоночника: А - миноги; Б - зародыша селахии; В - взрослой селахии; Г - рыбы. Схема позвонка: Д - миноги; Е, Ж - селахии; З - рыбы; И -рептилии; К-млекопитающего; 1 - передняя и 2 - задняя дорсальные дуги; 3 - миосепты; 4 - дорсальный и 5 - вентральный нервные корешки; 6 -хорда; 7- оболочка хорды; 8 - вентральные дуги; 9 - ребро; 10- сосудистые отростки; 11- поперечный отросток позвонка; 12 - тело позвонка; 13 - дужка позвонка; 14 - отверстие позвонка; 15 - остистый отросток; 16 - суставной отросток.
С усложнением организации животных, увеличением активности и разнообразия движений вокруг хорды развиваются не только дуги, но и тела позвонков, в результате она оказывается сдавленной, а у многих взрослых
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
форм и в различной степени редуцированной (Б, В, Г). Постепенное замещение хорды позвонками можно проследить, сравнивая строение осевого скелета у хрящевых и костистых рыб. У хрящевых рыб весь скелет образован хрящом, иногда обызвествленным. Кроме верхних дуг, также метамерно под хордой у них развиваются нижние дуги. Концы верхних дуг каждого сегмента, сливаясь, образуют остистый отросток. Появляются тела позвонков в виде двояковогнутых дисков (амфицельные) с отверстием в центре (Е, Ж). В канале, образованном отверстиями тел позвонков, проходит хорда, которая теперь теряет значение опорного стержня. Она принимает четкообразную форму с сужениями в телах позвонков и расширениями между позвонками. Появляются зачаточные ребра.
У некоторых видов хрящевых и костных (кистеперых, двоякодышащих, осетровых) рыб тела позвонков не развиваются, ребра отсутствуют. У всех костистых рыб происходит замещение хрящевого скелета костным, позвонки амфицельного типа с хорошо развитыми телами, дорсальными дугами и длинными остистыми отростками. Развиваются поперечные отростки, к которым причленяются хорошо развитые длинные ребра. Появляются суставные отростки, которыми позвонки сочленяются друг с другом, чем обеспечивается прочность осевого скелета при сохранении его подвижности. Осевой скелет четко разделяется на головной, туловищный с ребрами, охватывающими полость тела с органами, и сильно развитый хвостовой отдел - локомоторный.
Предки современных наземных позвоночных, по-видимому, произошли от древних кистеперых рыб, о чем говорит строение их ископаемых остатков. Переход к наземному образу жизни приводит к прогрессивному развитию одних частей скелета и редукции других. Скелет туловища дифференцируется на шейный, грудной (спинной), поясничный и крестцовый отделы, частично редуцируется скелет хвоста, так как основная нагрузка при движении по земле падает на конечности (кроме безногих амфибий и рептилий). В грудном отделе в тесной связи с ребрами развивается грудина, формируется грудная клетка.
Постепенность усложнения стволового скелета видна при сравнении классов наземных позвоночных. У земноводных шейный и крестцовый отделы позвоночника имеют только по одному позвонку, поясничный отдел отсутствует. Ребра очень короткие, у многих срастаются с поперечными отростками позвонков. Общее число позвонков сильно различается у амфибий разных отрядов: от 21 у бесхвостых до 300 у безногих. Тела их спереди вогнутые, сзади выпуклые (процельные). У грудины нет связи с ребрами, грудная клетка не образуется.
У рептилий шейный отдел удлиняется до восьми позвонков и приобретает большую подвижность. Два первых позвонка атлант и осевой (эпистрофей) сильно отличаются от остальных, к трем последним причленяются шейные ребра. Позвонки процельные (И). В грудном отделе 1-5 пар ребер соединены с грудиной - образуется грудная клетка. Поясничный отдел длинный, имеет ребра, величина которых уменьшается в каудальном направ-
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
лении. Крестцовый отдел образован двумя позвонками, хвостовой отдел длинный, хорошо развит.
Предками млекопитающих, по-видимому, были мелкие зверозубые пресмыкающиеся (величиной с крысу или кролика) пермского периода, которые наряду с крайне примитивными признаками имели ряд черт сходства с млекопитающими. У млекопитающих, независимо от образа жизни, постоянное число шейных позвонков, равное 7. Исключение составляют ламантин и двупалый ленивец, имеющие 6 шейных позвонков, а трехпалый ленивец - 8- 10. Относительно постоянно число позвонков и в остальных отделах: 12-19 грудных, 5-7 поясничных, 3-9 крестцовых. Хвостовых позвонков насчитывают от 3 до 46. Позвонки, за исключением первых двух, соединены при помощи хрящевых дисков (менисков), связок и суставных отростков.
Поверхности тел шейных позвонков часто имеют выпукло-вогнутую форму - опистоцельные (К). В остальных отделах позвонки обычно плоские
Платицельные.
Ребра сохраняются только в грудном отделе. В пояснице они редуцируются и срастаются с поперечными отростками позвонков. В крестцовом отделе срастаются и позвонки, образуя крестцовую кость. Хвостовой отдел облегчен, его позвонки сильно редуцируются.
Филогенез скелета головы (рис. 42). Скелет головного конца тела развивается вокруг нервной трубки - осевой (мозговой) скелет головы и вокруг головной кишки - висцеральный. У примитивных хордовых (круглоротые) они не связаны друг с другом. Осевой скелет головы представлен хрящевыми пластинками, окружающими нервную трубку снизу и с боков, крыша черепа перепончатая. Висцеральный скелет головы состоит из хрящевых жаберных дуг, связанных с аппаратом дыхания и пищеварения; челюстей нет.
Филогенетическое развитие скелета головы шло путем объединения мозгового и висцерального скелетов и усложнения их строения в связи с прогрессивным развитием головного мозга, органов чувств, расположенных на головном конце тела (обоняния, зрения, слуха), преобразованиями органов захвата и удержания пищи и дыхательного аппарата. Мозговой череп хрящевых рыб представляет собой сплошную хрящевую коробку, окружающую головной мозг. Висцеральный скелет образован хрящевыми жаберными дугами, которые подобно ребрам в туловище опоясывают собой головную часть кишечного канала. Передние висцеральные дуги превратились в губные хрящи, челюстную и подъязычную дуги.
Черепная коробка у костистых рыб сложного строения. Наряду с первичными костями, появившимися в большом количестве, на месте хрящевого черепа развиваются покровные кости. Первичные кости формируют затылочную область, часть основания черепа, обонятельную и слуховую капсулы и стенку глазницы. Покровные кости покрывают первичную черепную коробку сверху, снизу и с боков. Висцеральный скелет костистых рыб построен из большого количества первичных и вторичных костей и представляет собой очень сложную систему рычагов, участвующих в хватательных, глотательных и дыхательных движениях. С черепной коробкой висцеральный скелет

Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
сочленяется при помощи подвеска (hyomandibulare), в результате чего образуется единый скелет головы. С помощью костей плечевого пояса он неподвижно соединяется со стволовым скелетом.
Рис. 42. Филогенетические преобразования скелета головы (покровные кости белые, первичные кости и хрящи, из которых они развились, в
А-селахии; Б - костистой рыбы; В - млекопитающего: а - обонятельная, б - глазничная, в - слуховая и г - затылочная области черепа;1
Губные хрящи; 2 - нёбно-квадратный хрящ; 2" - квадратная кость; 2" - наковальня; 3- челюстной хрящ; 3" - сочленовная кость; 3" - молоточек; 4 - подъязычночелюстной хрящ; 4" - подвесок; 4" - стремечко; 5 - подъязычный хрящ; 5" - подъязычная; 6 - носовая; 7 - лобная; 8 - теменная; 9 - межтеменная; 10-затылочная; 11 - височная; 12 - клиновидная и 13
- слезная кости; 14 - сошник; 15 - крыловидная. 16-нижнечелюстная, 17
- нёбная; 18 - решетчатая; 19 - верхнечелюстная и 20 - резцовая кости; I-V - жаберные дуги; Г - тело подъязычной кости; 1Г-IIГ - щитовидный хрящ; IV-V - остальные хрящи гортани.
С выходом на сушу, с резкой сменой среды обитания, а следовательно, и образа жизни животных происходят значительные изменения в скелете головы. Череп теряет связь с плечевым поясом и подвижно причленяется к шей-
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
ному отделу. Уменьшается количество костей черепа за счет их сращения, увеличивается его прочность. Смена типа дыхания (с жаберного на легочное) приводит к редукции жаберного аппарата и преобразованию его элементов, а также некоторых покровных костей висцерального скелета, в подъязычную и слуховые кости.
Преобразования в строении черепа приводят к срастанию челюстного аппарата с основанием черепа, к появлению слуховых косточек в обособившейся полости среднего уха, к объединению обонятельной капсулы с носовой полостью, к разделению носовой и ротовой полостей с помощью твердого нёба и изменению характера причленения нижней челюсти к осевому черепу.
В ряду наземных хордовых животных прослеживается постепенность этих усложнений. В черепе взрослых земноводных много хрящей, слуховая кость одна - стремечко (столбик). По числу костей черепа рептилии скорее приближаются к рыбам, чем к амфибиям, однако строение черепа типично для наземных животных. Для черепа млекопитающих характерно прогрессивное развитие, выраженное в уменьшении количества костей за счет их сращения (например, затылочная кость образуется путем сращения 4, а каменистая- 5 костей), в стирании граней между первичными и покровными (вторичными) костями, в мощном развитии обонятельной области и сложном звукопроводящем аппарате, в крупных размерах черепной коробки, резко выраженной разнозубости, самостоятельном причленении нижней челюсти к слуховой области черепа (без каких-либо промежуточных костей).
Филогенез скелета конечностей (рис. 43). Гипотеза о происхождении конечностей наземных животных на базе парных плавников рыб оказалась плодотворной и сейчас имеет широкое признание. Парные плавники в типе хордовых впервые появились у рыб (Л). У круглоротых они отсутствуют, есть, как и у ланцетника, только непарные плавники. Костной основой парных плавников рыб служит целая система хрящевых и костных элементов, которые могут быть подразделены на несколько отделов. Самый проксимальный отдел, называемый поясом грудных плавников, у хрящевых рыб имеет вид изогнутой пластинки, прилежащей к осевому скелету и соединяющей правый и левый плавники в единую систему. У костистых рыб он более сложной формы и прочнее связан с осевым скелетом. Среди его костей есть две небольшие кости, гомологичные костям плечевого пояса наземных животных: лопатка - лежащая на спинной стороне и коракоид - на брюшной.
Тазовый пояс у рыб развит в меньшей степени. Имеет вид хрящевой или костной пластинки, лежащей в мускулатуре брюшной стенки. К его боковой поверхности причленяется скелет брюшного плавника. У современных лучеперых рыб плавники претерпели редукцию и видоизменение в сравнении с костными гоноидами и двудышащими. По строению к конечностям наземных позвоночных ближе всего стоят плавники и их пояса у ископаемых кистеперых рыб (Б).

Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
Рис. 43. Филогенетическое преобразование скелета конечностей:
А -парный грудной плавник селахии; Б - грудной плавник кистеперой рыбы; В - скелет грудной конечности примитивного наземного позвоночного; Г - скелет грудной конечности стопоходящего млекопитающего (медведя); Д - изменения кисти у предков лошади: а - эогиппус; б - мезогиппус; в - гиппогиппус; г - гиппарион; д - современная лошадь; 1-плечевой пояс: 2 - базалии; 3- радиалии; 4 - лопатка; 5 - ключица; 6 - коракоидная кость; 7 - плечевая кость; 8 - кости предплечья; 9 - кости кисти; 10- кости запястья; 11 - кости пясти; 12 - кости пальцев.
С выходом на сушу полностью исчезает скелет непарных плавников. На базе парных плавников развивается скелет конечностей, расчлененный на отделы, типичные для пятипалой конечности (В). Пояса конечностей состоят из трех пар костей и укрепляются связью с осевым скелетом: плечевой пояс - с грудиной, тазовый с крестцом. Плечевой пояс состоит из коракоида, лопатки и ключицы, тазовый - из подвздошной, лонной и седалищной костей. Скелет свободных конечностей расчленен на три отдела: у передней конечности
Это кости плеча, предплечья и кисти, у задней - бедра, голени и стопы. Число костей в гомодинамных звеньях конечностей одинаково и увеличивается в дистальном направлении.
Дальнейшие филогенетические преобразования связаны с характером передвижения, его скоростью и маневренностью. У земноводных пояс грудных конечностей, причленяясь к грудине, не имеет жесткой связи с осевым скелетом. В поясе тазовых конечностей развита в основном его вентральная часть. Причленяется он к единственному крестцовому позвонку. Свободные конечности присоединяются к поясам в сегментальной плоскости, так что плечо и бедро отходит от тела под прямым углом, а лапы направлены латеро-
Вракин В.Ф, Сидорова М.В. |
МОРФОЛОГИЯ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ |
каудально. У рептилий в скелете поясов в равной степени развиты дорсальная и вентральная части. Свободные конечности отходят от тела, как и у амфибий, под прямым углом, но их дистальные звенья развернуты, особенно на грудной конечности, в латеро-краниальном направлении.
Плечевой пояс млекопитающих (Г) сильно редуцирован, особенно его вентральное звено. Он состоит из двух или даже одной кости. У животных с развитыми отводящими движениями грудной конечности (например, кротов, летучих мышей, обезьян) развиты лопатка и ключица, а у животных с однообразными движениями (например, у копытных)-только лопатка. Тазовый пояс млекопитающих упрочен тем, что лонные и седалищные кости соединяются вентрально с соименными костями. Скелет свободных конечностей млекопитающих развернут в сагиттальной плоскости, лапы направлены краниально. Построен он по типу пятипалой конечности, с длинными проксимальными звеньями, в результате чего тело животного высоко поднято над землей. Приспособление же к различным видам передвижения (бег, лазанье, прыжки, полет, плавание) привело к сильной специализации конечностей у разных групп млекопитающих, что выражается в основном в изменении длины и угла наклона отдельных звеньев конечностей, формы суставных поверхностей, срастании костей и редукции пальцев.
Изменение в строении конечностей в филогенезе в связи с повышением специализации - приспособленностью к определенному виду движения подробнее всего изучено в ряду лошадей (В. О. Ковалевский). Предполагаемый предок лошади - эупротогония, совмещая в себе черты копытных и хищных, была размером с лисицу и имела пятипалые конечности с когтями, по форме приближающимися к копытцам. На протяжении нескольких периодов (от нижнего эоцена до нижнего плиоцена) происходила постепенная смена условий существования и параллельно этому шло приспособительное изменение структуры конечностей, связанное с изменением характера движения (Д). От разнообразных мягких движений по рыхлой земле с высокой растительностью (эоценовый лес) до широких размашистых быстрых движений по сухим открытым пространствам (миоценовая степь). При этом происходило удлинение основного поддерживающего столба конечностей из-за раскрытия (увеличения) углов между ее звеньями. Лапа приподнималась, животное переходило от стопо- к пальцехождению (эогиппус - четырехпалый, мезогип- пус-трехпалый). При этом наблюдалась постепенная редукция нефункционирующих пальцев: первого у эогиппуса, первого и пятого у мезогиппуса. При переходе от пальце- к фаланго-(копыто-) хождению в состав основного поддерживающего столба входит уже вся лапа, а редукция пальцев достигает максимума. У лошади полноразвитым сохраняется на конечности лишь III палец. У рогатого скота развитыми оказываются два пальца - III и IV.
ЛЕКЦИЯ МИОЛОГИЯ ФИЛОГЕНЕЗ, ОНТОГЕНЕЗ И ФУНКЦИОНАЛЬНАЯ АНАТОМИЯ МЫШЕЧНОЙ СИСТЕМЫ Выполнили: Владимирова Я. Б. Кокорева Т. В.

 Мышцы или мускулы (от лат. musculus - мышка, маленькая мышь) - органы тела животных и человека, состоящие из упругой, эластичной мышечной ткани, способной сокращаться под влиянием нервных импульсов. Предназначены для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 86, 3 % из воды. В теле человека 640 мышц
Мышцы или мускулы (от лат. musculus - мышка, маленькая мышь) - органы тела животных и человека, состоящие из упругой, эластичной мышечной ткани, способной сокращаться под влиянием нервных импульсов. Предназначены для выполнения различных действий: движения тела, сокращения голосовых связок, дыхания. Мышцы состоят на 86, 3 % из воды. В теле человека 640 мышц
 Мотивация: - - - возможности совершаемого движения, объём движения; активные или пассивные движения запускаются в действие той или иной мышечной группой; воздействуя на мышечный аппарат, мы изменяем общее состояние; мышечный рельеф является ориентиром для топографии сосудов и нервов; пересадка мышц, то есть мышца может «переучиваться» .
Мотивация: - - - возможности совершаемого движения, объём движения; активные или пассивные движения запускаются в действие той или иной мышечной группой; воздействуя на мышечный аппарат, мы изменяем общее состояние; мышечный рельеф является ориентиром для топографии сосудов и нервов; пересадка мышц, то есть мышца может «переучиваться» .
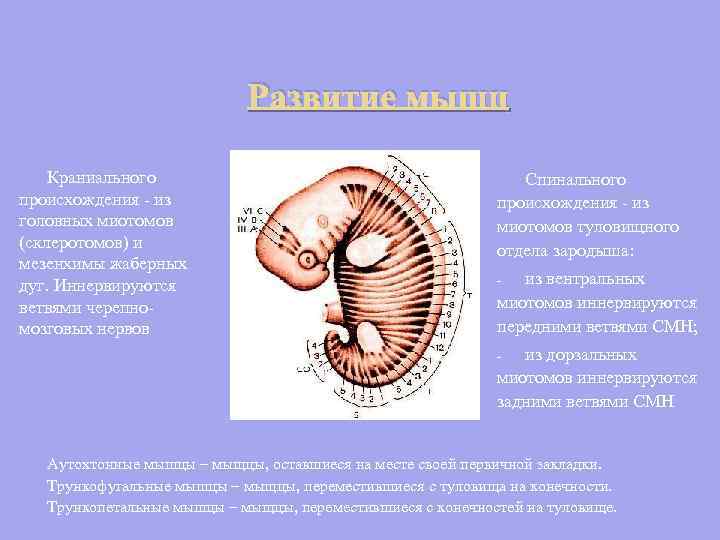 Развитие мышц Краниального происхождения - из головных миотомов (склеротомов) и мезенхимы жаберных дуг. Иннервируются ветвями черепномозговых нервов Спинального происхождения - из миотомов туловищного отдела зародыша: из вентральных миотомов иннервируются передними ветвями СМН; - из дорзальных миотомов иннервируются задними ветвями СМН - Аутохтонные мышцы – мыщцы, оставшиеся на месте своей первичной закладки. Трункофугальные мышцы – мыщцы, переместившиеся с туловища на конечности. Трункопетальные мышцы – мыщцы, переместившиеся с конечностей на туловище.
Развитие мышц Краниального происхождения - из головных миотомов (склеротомов) и мезенхимы жаберных дуг. Иннервируются ветвями черепномозговых нервов Спинального происхождения - из миотомов туловищного отдела зародыша: из вентральных миотомов иннервируются передними ветвями СМН; - из дорзальных миотомов иннервируются задними ветвями СМН - Аутохтонные мышцы – мыщцы, оставшиеся на месте своей первичной закладки. Трункофугальные мышцы – мыщцы, переместившиеся с туловища на конечности. Трункопетальные мышцы – мыщцы, переместившиеся с конечностей на туловище.



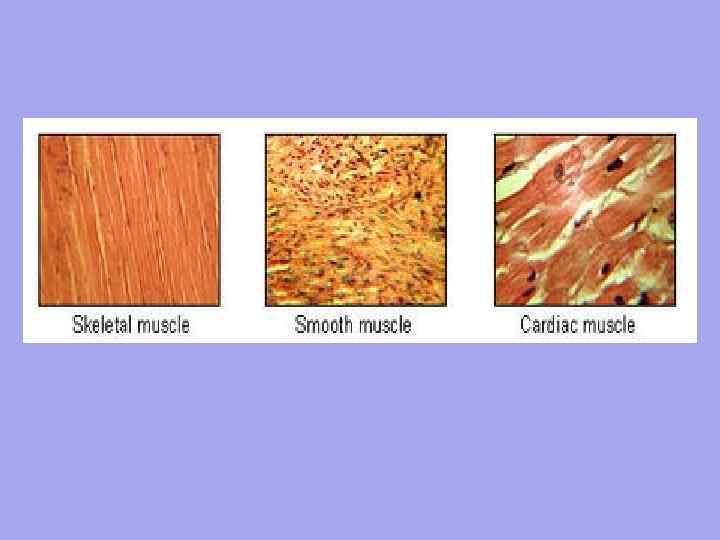
 Поперечно-полосатая Гладкая 1. Единицей организации является миоцит. Длина около 50 мкм. Ширина от 6 мкм. 2. Непроизвольное сокращение Контроль со стороны автономной нервной системы Движение волнообразное, работает медленно, так как нервное волокно подходит НЕ к каждой клетке Медленно вступают в действие, но сохраняются на длительном протяжении Не имеет точной пространственной ориентации клеток 3. 4. 5. 6. 1. 2. 3. 4. 5. 6. Сердечная Единицей организации является мышечное волокно- множество миобластов, плавающих в общей цитоплазме (саркоплазме). У них общая сарколемма. Длина около 40 -100 мм. Ширина от 7 мм. Произвольное сокращение Контроль со стороны соматической нервной системы Быстрое сокращение, быстрая реакция, так каждое мышечное волокно имеет нервно -мышечный синапс Быстро включаются, но имеет кратковременное действие Четкая ориентация мышечных волокон
Поперечно-полосатая Гладкая 1. Единицей организации является миоцит. Длина около 50 мкм. Ширина от 6 мкм. 2. Непроизвольное сокращение Контроль со стороны автономной нервной системы Движение волнообразное, работает медленно, так как нервное волокно подходит НЕ к каждой клетке Медленно вступают в действие, но сохраняются на длительном протяжении Не имеет точной пространственной ориентации клеток 3. 4. 5. 6. 1. 2. 3. 4. 5. 6. Сердечная Единицей организации является мышечное волокно- множество миобластов, плавающих в общей цитоплазме (саркоплазме). У них общая сарколемма. Длина около 40 -100 мм. Ширина от 7 мм. Произвольное сокращение Контроль со стороны соматической нервной системы Быстрое сокращение, быстрая реакция, так каждое мышечное волокно имеет нервно -мышечный синапс Быстро включаются, но имеет кратковременное действие Четкая ориентация мышечных волокон
 Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани- эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него, что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по нескольку мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием.
Между мышечными волокнами находятся тонкие прослойки рыхлой волокнистой соединительной ткани- эндомизий. Коллагеновые волокна наружного листка базальной мембраны вплетаются в него, что способствует объединению усилий при сокращении миосимпластов. Более толстые прослойки рыхлой соединительной ткани окружают по нескольку мышечных волокон, образуя перимизий и разделяя мышцу на пучки. Несколько пучков объединяются в более крупные группы, разделенные более толстыми соединительнотканными прослойками. Соединительную ткань, окружающую поверхность мышцы, называют эпимизием.

 В мышце как органе есть соединительная ткань Эндомизий – тонкая соединительная ткань, окружает каждое мышечное волокно и небольшие группы волокон. Перимизий – покрывает более крупные комплексы мышечных волокон и мышечные пучки.
В мышце как органе есть соединительная ткань Эндомизий – тонкая соединительная ткань, окружает каждое мышечное волокно и небольшие группы волокон. Перимизий – покрывает более крупные комплексы мышечных волокон и мышечные пучки.
 Значение эндомизия и перимизия 1. Через эндомизиум и перимизиум к мышечному волокну подходят сосуды и нервы. Образуют строму органа; 2. Мышечные волокна формируются в пучки, пучки в мышцу; 3. Так как эндомизий сращен с сарколеммой мышечного волокна, следовательно, сокращающее мышечное волокно может растягиваться лишь до определенного предела
Значение эндомизия и перимизия 1. Через эндомизиум и перимизиум к мышечному волокну подходят сосуды и нервы. Образуют строму органа; 2. Мышечные волокна формируются в пучки, пучки в мышцу; 3. Так как эндомизий сращен с сарколеммой мышечного волокна, следовательно, сокращающее мышечное волокно может растягиваться лишь до определенного предела
 Миофибриллы в волокне окружены оболочкой – сарколеммой, и погружены в специальную среду – саркоплазму. В зависимости от содержания пигмента и кислорода волокна делятся на белые и красные. Белые волокна –анаэробы, содержат больше миофибрилл, меньше саркоплазмы. Запускаются быстро, но долго работать не могут. Пример: грудинно-ключичнососцевидная, икроножная мышцы. Красные волокна – волокна толстые. Много миоглобина в саркоплазме и цитохрома в митохондриях, но меньше миофибрилл. Медленно запускаются, но работают долго. Пример: мышцы спины, диафрагма.
Миофибриллы в волокне окружены оболочкой – сарколеммой, и погружены в специальную среду – саркоплазму. В зависимости от содержания пигмента и кислорода волокна делятся на белые и красные. Белые волокна –анаэробы, содержат больше миофибрилл, меньше саркоплазмы. Запускаются быстро, но долго работать не могут. Пример: грудинно-ключичнососцевидная, икроножная мышцы. Красные волокна – волокна толстые. Много миоглобина в саркоплазме и цитохрома в митохондриях, но меньше миофибрилл. Медленно запускаются, но работают долго. Пример: мышцы спины, диафрагма.
 Каждая мышца имеет сеть кровеносных сосудов. Сокращения мышцы способствует току крови. В расслабленной неработающей мышце большая часть кровеносных капилляров закрыта для тока крови. При сокращении мышцы все кровеносные капилляры сразу открываются.
Каждая мышца имеет сеть кровеносных сосудов. Сокращения мышцы способствует току крови. В расслабленной неработающей мышце большая часть кровеносных капилляров закрыта для тока крови. При сокращении мышцы все кровеносные капилляры сразу открываются.

 Строение мышцы Каждая мышца одним концом соединяется с одной костью (начало мышцы), а другим – с другой (прикрепление мышцы). В мышце различают: головку брюшко хвостовую часть.
Строение мышцы Каждая мышца одним концом соединяется с одной костью (начало мышцы), а другим – с другой (прикрепление мышцы). В мышце различают: головку брюшко хвостовую часть.
 К каждому мышечному волокну подходят двигательные нервные волокна и отходят чувствительные нервные волокна Количество нервных окончаний в мышце зависит от степени функциональной активности мышц.
К каждому мышечному волокну подходят двигательные нервные волокна и отходят чувствительные нервные волокна Количество нервных окончаний в мышце зависит от степени функциональной активности мышц.

 Каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом. Группа же мышечных волокон, иннервируемых одним мотонейроном, называется моторной единицей. Характерно, что мышечные волокна, принадлежащие к одной моторной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.
Каждое мышечное волокно иннервируется самостоятельно и окружено сетью гемокапилляров, образуя комплекс, именуемый мионом. Группа же мышечных волокон, иннервируемых одним мотонейроном, называется моторной единицей. Характерно, что мышечные волокна, принадлежащие к одной моторной единице, лежат не рядом, а расположены мозаично среди волокон, относящихся к другим единицам.


 Сухожилие – это плотный волокнистый соединительно-тканный тяж, которым мышца начинается или прикрепляется к скелету
Сухожилие – это плотный волокнистый соединительно-тканный тяж, которым мышца начинается или прикрепляется к скелету
 перитенониум коллагеновые волокна IV типа эндотенониум Коллагеновые волокна сухожилия, переплетающиеся с коллагеновыми волокнами надкостницы, вплетаются в основное вещество костной ткани, образуя на костях гребни, бугорки, бугристости, впадины, вдавления.
перитенониум коллагеновые волокна IV типа эндотенониум Коллагеновые волокна сухожилия, переплетающиеся с коллагеновыми волокнами надкостницы, вплетаются в основное вещество костной ткани, образуя на костях гребни, бугорки, бугристости, впадины, вдавления.
 Фасции – это соединительнотканные коллагенные волокна с небольшой примесью эластических волокон Поверхностная височная фасция Глубокие фасции бедра
Фасции – это соединительнотканные коллагенные волокна с небольшой примесью эластических волокон Поверхностная височная фасция Глубокие фасции бедра
 1. 2. 3. 4. 5. Фасции отделяют мышцы от кожи и устраняют смещение кожи при движениях сокращающихся мышц. Фасции экономят силу сокращения мышц, устраняя трение между мышцами во время сокращения. Фасции растягивают крупные вены при натяжении, в результате чего кровь с периферии «присасывается» в эти вены. Фасции имеют значение как барьеры, препятствующие распространению инфекции и опухолей. Во время операций фасции помогают определять расположение мышц, кровеносных сосудов, внутренностей.
1. 2. 3. 4. 5. Фасции отделяют мышцы от кожи и устраняют смещение кожи при движениях сокращающихся мышц. Фасции экономят силу сокращения мышц, устраняя трение между мышцами во время сокращения. Фасции растягивают крупные вены при натяжении, в результате чего кровь с периферии «присасывается» в эти вены. Фасции имеют значение как барьеры, препятствующие распространению инфекции и опухолей. Во время операций фасции помогают определять расположение мышц, кровеносных сосудов, внутренностей.
 Классификация мышц Скелетные мышцы разнообразны по форме, структуре, положению относительно осей суставов и т. д. , поэтому классифицируются по-разному.
Классификация мышц Скелетные мышцы разнообразны по форме, структуре, положению относительно осей суставов и т. д. , поэтому классифицируются по-разному.

 III. По функциональной особенности Статические (сильные) – короткое брюшко и длинное сухожилие. Мышцы работают с большей силой, но с меньшим размахом движения. Динамические (ловкие) – длинные мышечные пучки, сухожилия короткие. Мышцы работают с меньшей силой, но производят движения большего размаха
III. По функциональной особенности Статические (сильные) – короткое брюшко и длинное сухожилие. Мышцы работают с большей силой, но с меньшим размахом движения. Динамические (ловкие) – длинные мышечные пучки, сухожилия короткие. Мышцы работают с меньшей силой, но производят движения большего размаха
 Вспомогательный аппарат мышц Скелетные мышцы имеют вспомогательный аппарат, облегчающий их функционирование. n n n Фасции; Костно-фасциальные влагалища; Синовиальные сумки; Синовиальные влагалища сухожилий; Мышечные блоки; Сесамовидные кости.
Вспомогательный аппарат мышц Скелетные мышцы имеют вспомогательный аппарат, облегчающий их функционирование. n n n Фасции; Костно-фасциальные влагалища; Синовиальные сумки; Синовиальные влагалища сухожилий; Мышечные блоки; Сесамовидные кости.

 Аномалии развития мышц Встречаются очень часто и делятся на три группы: 1. Отсутствие какой-либо мышцы; 2. Наличие дополнительной мышцы, которой нет в природе. 3. Дополнительные пучки имеющейся мышцы.
Аномалии развития мышц Встречаются очень часто и делятся на три группы: 1. Отсутствие какой-либо мышцы; 2. Наличие дополнительной мышцы, которой нет в природе. 3. Дополнительные пучки имеющейся мышцы.
 Пороки развития Недоразвитие грудинно-ключично-сосцевидной мышцы – Кривошея Недоразвитие диафрагмы. Причина появление диафрагмальных грыж. Недоразвитие дельтовидной и трапециевидной мышц – Деформация плечевого пояса и плеча
Пороки развития Недоразвитие грудинно-ключично-сосцевидной мышцы – Кривошея Недоразвитие диафрагмы. Причина появление диафрагмальных грыж. Недоразвитие дельтовидной и трапециевидной мышц – Деформация плечевого пояса и плеча


 I. По форме: Веретенообразные; Лентовидные; Плоские широкие; Зубчатые; Длинные; n n n Квадратные; Треуголные; Круглые; Дельтовидная; Камбаловидная и т. д.
I. По форме: Веретенообразные; Лентовидные; Плоские широкие; Зубчатые; Длинные; n n n Квадратные; Треуголные; Круглые; Дельтовидная; Камбаловидная и т. д.



 II. По направлению мышечных волокон С прямыми параллельными волокнами; С поперечными; С круговыми; Перистые: A. Одноперистые; Двуперистые; C. Многоперистые. B.
II. По направлению мышечных волокон С прямыми параллельными волокнами; С поперечными; С круговыми; Перистые: A. Одноперистые; Двуперистые; C. Многоперистые. B.

 IV. По функции: Приводящие; Отводящие; Сгибающие; Разгибающие; Пронаторы; n n Супинаторы; Напрягающие; Мышцы – синергисты; Мышцы – антагонисты.
IV. По функции: Приводящие; Отводящие; Сгибающие; Разгибающие; Пронаторы; n n Супинаторы; Напрягающие; Мышцы – синергисты; Мышцы – антагонисты.
 V. По отношению к суставу: Односуставные; Двусуставные; Многосуставные.
V. По отношению к суставу: Односуставные; Двусуставные; Многосуставные.
Филогенез систем органов хордовых рассмотрен в соответствии с прогрессивным направлением эволюции этого типа животных от подтипа Бесчерепные до класса Млекопитающие. Организация систем органов класса Птицы не описана в связи с тем, что птицы произошли от пресмыкающихся значительно позже млекопитающих и являются боковой ветвью эволюции хордовых.
Наружные покровы
Покровы любых животных всегда выполняют функцию восприятия внешних раздражении, а также защищают тело от вредных воздействий среды. Интенсификация первой функции покровов приводит в процессе эволюции многоклеточных животных к возникновению нервной системы и органов чувств. Интенсификация второй функции сопровождается дифференцировкой. Характерно также расширение функций, в результате чего кожа как орган защиты участвует также в газообмене, терморегуляции и выделении, выкармливании потомства. Это связано с усложнением строения слоев кожи, появлением и дальнейшим преобразованием многочисленных придатков и желез.
У всех хордовых кожа имеет двойное - экто- и мезодермальное - происхождение. Из эктодермы развивается эпидермис, из мезодермы - дерма. Для бесчерепных характерна слабая степень дифференцированности обоих слоев кожи. Эпидермис однослойный цилиндрический, содержащий одноклеточные слизистые железы, дерма рыхлая, содержит небольшое количество соединительнотканных клеток.
В подтипе Позвоночные эпидермис становится многослойным, причем в нижнем слое клетки постоянно размножаются, а в верхних слоях - дифференцируются, гибнут и слущиваются. В дерме появляются соединительнотканные волокна, придающие покровам прочность. Кожа образует придатки, многообразные в зависимости от образа жизни и уровня организации, а также железы, выполняющие различные функции.
У рыб в эпидермисе железы одноклеточные. Как и у ланцетника, они выделяют слизь, облегчающую движения в воде. Тело рыб покрыто чешуей, имеющей разное строение в зависимости от их систематического положения. Чешую хрящевых рыб называют плакоидной. Она имеет форму шипа и состоит из дентина, покрытого снаружи эмалью (рис. 14.1). Дентин мезодермального происхождения, он образуется за счет функционирования соединительнотканных клеток, выпячивающихся снаружи в виде сосочка. Эмаль, представляющая собой более твердое, чем дентин, неклеточное вещество, формируется сосочком эпидермиса и покрывает плакоидную чешую снаружи.
Вся поверхность тела хрящевых рыб, а также ротовая полость, слизистая оболочка которой происходит из эктодермы, покрыты плакоидной чешуей. Естественно, что функции чешуи в ротовой полости связаны с захватом и удержанием пищи, поэтому они сильно увеличены и являются зубами. У костных рыб чешуя иного типа. Она имеет вид тонких круглых костных пластинок, покрытых тонким слоем эпидермиса. Костная чешуя развивается полностью за счет дермы, но по происхождению связана с примитивной плакоидной.
Кожа примитивных вымерших земноводных - стегоцефалов - соответствовала покровам рыб и тоже была покрыта чешуей. Современные амфибии имеют тонкую гладкую кожу без чешуи, принимающую участие в газообмене. Этому способствует наличие большого количества многоклеточных слизистых желез, секрет которых постоянно увлажняет покровы и обладает бактерицидными свойствами. Некоторые кожные железы ряда земноводных дифференцировались в органы-продуценты токсинов, защищающих их от врагов (см. разд. 23.1).
Рис. 14.1. Закладка плакоидной чешуи:
1 -клетки-эмалеобразователи, 2- эпидермис, 3- эмаль, 4- склеробласты-дентинообразователи, 5- дентин, 6- сосочек дермы
Пресмыкающиеся, перешедшие полностью к наземному существованию, имеют сухую кожу, не участвующую в дыхании. Верхний слой эпидермиса ороговевает. Роговые чешуйки у некоторых рептилий тонки и эластичны, у других-сливаются вместе, образуя, как у черепах, мощный роговой панцирь. Большинство пресмыкающихся по мере роста линяют, сбрасывая периодически свой роговой покров. Современные пресмыкающиеся не имеют кожных желез.
Кожные покровы млекопитающих построены наиболее сложно в связи с выполнением ими многообразных функций. Характерны различные производные кожи: волосы, когти, рога, копыта, а также потовые, сальные и млечные железы. Более примитивные млекопитающие - насекомоядные, грызуны и некоторые другие - наряду с волосяным покровом сохранили также роговые чешуи на хвосте. Волосы у них растут в промежутках между чешуйками, группами по 3-7. У более прогрессивных млекопитающих, утративших чешуи, сохраняется такое же расположение волос (рис. 14.2), покрывающих практически все тело, кроме некоторых участков, например подошв и ладоней у человека.
Волосы многих млекопитающих дифференцированы на типичные, служащие для терморегуляции, и большие, или вибриссы , основания которых связаны с чувствительными нервными окончаниями. У большинства млекопитающих вибриссы расположены в области рта и носа, у приматов они редуцированы в связи с усилением осязательной функции передних конечностей, у многих яйцекладущих и сумчатых - разбросаны по всему телу. Этот факт может свидетельствовать о том, что волосяной покров предков млекопитающих первично выполнял осязательные функции, а затем, по мере увеличения количества волос, стал принимать участие в терморегуляции. В онтогенезе человека закладывается большее количество волосяных зачатков, но к концу эмбриогенеза наступает редукция большинства из них.
Потовые железы млекопитающих гомологичны кожным железам амфибий. Их секрет может быть слизистым, содержать белки и жир. Некоторые потовые железы дифференцировались у ранних млекопитающих в млечные железы. У яйцекладущих (утконос, ехидна) млечные железы сходны с потовыми по строению и развитию. По краям развивающегося соска молочной железы можно обнаружить последовательные переходы от типичных потовых к млечным железам (рис. 14.3). Количество млечных желез и сосков коррелирует с плодовитостью (от 25 до одной пары), но в эмбриогенезе всех млекопитающих на брюшной поверхности закладываются «млечные линии», тянущиеся от подмышечной впадины до паха. Впоследствии на этих линиях дифференцируются соски, большая часть которых затем подвергается редукции и исчезает. Так, в эмбриогенезе человека закладывается вначале пять пар сосков, а впоследствии остается лишь одна (рис. 14.4).

Рис. 14.4. Эмбриогенез передней брюшной стенки человека. А - зародыш в возрасте 5 недель (видны млечные линии); Б - дифференцировка пяти пар сосков; В - зародыш в возрасте 7 недель

Рис. 14.5. Атавистические аномалии развития кожи.
А - гипертрихоз; Б - полимастия
Сальные железы образуются в коже только у млекопитающих. Их секрет, смазывая волосы и поверхность кожи, придает им несмачиваемость и эластичность.
Онтогенез покровов и придатков кожи млекопитающих и человека отражает их эволюцию по типу архаллаксиса. Действительно, ни зачатки роговых чешуи, характерных для пресмыкающихся, ни более ранние формы придатков кожи в их эмбриогенезе не рекапитулируют. При этом на стадии вторичного органогенеза развиваются сразу зачатки волосяных фолликулов. Нарушения раннего онтогенеза кожных покровов человека могут вызвать возникновение некоторых малосущественных атавистических пороков развития: гипертрихоз (повышенное оволосение), полителию (увеличенное количество сосков), полимастию (увеличенное количество млечных желез) (рис. 14.5). Все они связаны с нарушением редукции избыточного количества этих структур и отражают эволюционную связь человека с наиболее близкими предковыми формами - млекопитающими. Именно поэтому у человека и других млекопитающих невозможно рождение потомства с атавистическими признаками кожных покровов, характерными для более отдаленных предков. Одним из самых известных признаков недоношенности новорожденных является повышенное оволосение кожи. Вскоре после рождения избыточные волосы обычно выпадают, а их фолликулы редуцируются.
ОПОРНО-ДВИГАТЕЛЬНЫЙ АППАРАТ
Филогенез двигательной функции лежит в основе прогрессивной эволюции животных. Поэтому уровень их организации в первую очередь зависит от характера двигательной активности, которая определяется особенностями организации опорно-двигательного аппарата, претерпевшего в типе Хордовые большие эволюционные преобразования в связи со сменой сред обитания и изменения форм локомоции. Действительно, водная среда у животных, не имеющих наружного скелета, предполагает однообразные движения за счет изгибов всего тела, в то время как жизнь на суше более способствует их перемещению с помощью конечностей.
Рассмотрим в отдельности эволюцию скелета и мышечной системы.
Скелет
У хордовых скелет внутренний. По строению и функциям подразделяется на осевой, скелет конечностей и головы.
Осевой скелет
В подтипе Бесчерепные имеется только осевой скелет в виде хорды. Она построена из сильно вакуолизированных клеток, плотно прилегающих друг к другу и покрытых снаружи общими эластической и волокнистой оболочками. Упругость хорде придают тургорное давление ее клеток и прочность оболочек. Хорда закладывается в онтогенезе всех хордовых и выполняет у более высокоорганизованных животных не столько опорную, сколько морфогенетическую функцию, являясь органом, осуществляющим эмбриональную индукцию.
На протяжении всей жизни у позвоночных хорда сохраняется только у круглоротых и некоторых низших рыб. У всех остальных животных она редуцируется. У человека в постэмбриональном периоде сохраняются рудименты хорды в виде nucleus pulposus межпозвоночных дисков. Сохранение избыточного количества хордального материала при нарушении его редукции чревато возможностью развития у человека опухолей - хордом, возникающих на его основе.
У всех позвоночных хорда постепенно вытесняется позвонками, развивающимися из склеротомов сомитов, и функционально заменяется позвоночным столбом. Это один из выраженных примеров гомотопной субституции органов (см. § 13.4). Формирование позвонков в филогенезе начинается с развития их дуг, охватывающих нервную трубку и становящихся местами прикрепления мышц. Начиная с хрящевых рыб обнаруживается охрящевение оболочки хорды и разрастание оснований позвонковых дуг, в результате чего формируются тела позвонков. Срастание верхних позвонковых дуг над нервной трубкой образует остистые отростки и позвоночный канал, в который заключена нервная трубка (рис. 14.6).

Рис. 14.6. Развитие позвонка. А-ранний этап; Б- последующая стадия:
1 -хорда, 2- оболочка хорды, 3- верхние и нижние позвонковые дуги, 4- остистый отросток, 5- зоны окостенения, 6-рудимент хорды, 7- хрящевое тело позвонка
Замещение хорды позвоночным столбом - более мощным органом опоры, имеющим сегментарное строение,- позволяет увеличить общие размеры тела и активизирует двигательную функцию. Дальнейшие прогрессивные изменения позвоночного столба связаны с тканевой субституцией - заменой хрящевой ткани на костную, что обнаруживается у костных рыб, а также с дифференцировкой его на отделы.
У рыб только два отдела позвоночника: туловищный и хвостовой. Это связано с перемещением их в воде за счет изгибов тела.
Земноводные приобретают также шейный и крестцовый отделы, представленные каждый одним позвонком. Первый обеспечивает большую подвижность головы, а второй - опору задним конечностям.
У пресмыкающихся удлиняется шейный отдел позвоночника, первые два позвонка которого подвижно соединены с черепом и обеспечивают большую подвижность головы. Появляется поясничный отдел, еще слабо отграниченный от грудного, а крестец состоит уже из двух позвонков.
Млекопитающие характеризуются стабильным количеством позвонков в шейном отделе, равным 7. В связи с большим значением в движении задних конечностей крестец образован 5-10 позвонками. Поясничный и грудной отделы четко отграничены друг от друга.
У рыб все туловищные позвонки несут ребра, не срастающиеся друг с другом и с грудиной. Они придают телу устойчивую форму и обеспечивают опору мышцам, изгибающим тело в горизонтальной плоскости. Эта функция ребер сохраняется у всех позвоночных, совершающих змеевидные движения,- у хвостатых земноводных и пресмыкающихся, поэтому у них ребра также располагаются на всех позвонках, кроме хвостовых.
У пресмыкающихся часть ребер грудного отдела срастается с грудиной, формируя грудную клетку, а у млекопитающих в состав грудной клетки входит 12-13 пар ребер.

Рис. 14.7. Аномалии развития осевого скелета. А - рудиментарные шейные ребра (показаны стрелками); Б - несращение остистых отростков позвонков в грудной и поясничной областях. Спинномозговые грыжи
Онтогенез осевого скелета человека рекапитулирует основные филогенетические стадии его становления: в периоде нейруляции закладывается хорда, заменяющаяся впоследствии хрящевым, а затем и костным позвоночником. На шейных, грудных и поясничных позвонках развивается по паре ребер, после чего шейные и поясничные ребра редуцируются, а грудные срастаются спереди друг с другом и с грудиной, формируя грудную клетку.
Нарушение онтогенеза осевого скелета у человека может выразиться в таких атавистических пороках развития, как несрастание остистых отростков позвонков, в результате чего формируется spinabifida - дефект позвоночного канала. При этом часто через дефект выпячиваются мозговые оболочки и образуется спинномозговая грыжа (рис. 14.7).
В возрасте 1,5-3 мес. зародыш человека обладает хвостовым отделом позвоночника, состоящим из 8-11 позвонков. Нарушение их редукции в последующем объясняет возможность возникновения такой известной аномалии осевого скелета, как персистирование хвоста.
Нарушение редукции шейных и поясничных ребер лежит в основе их сохранения в постнатальном онтогенезе.
Скелет головы
Продолжением осевого скелета спереди является осевой, или мозговой, череп, служащий для защиты головного мозга и органов чувств. Рядом с ним развивается висцеральный, или лицевой череп, образующий опору передней части пищеварительной трубки. Обе части черепа развиваются по-разному и из разных зачатков. На ранних этапах эволюции и онтогенеза они не связаны между собой, но позже эта связь возникает.

Рис. 14.8. Череп человека с методическим швом (указан стрелкой)
В задней части осевого черепа в процессе развития обнаруживаются следы сегментации, поэтому считают, что он представляет собой результат слияния друг с другом закладок передних позвонков. В состав мозгового черепа включаются также закладки хрящевых капсул мезенхимального происхождения, окружающие органы слуха, обоняния и зрения. Кроме того, часть мозгового черепа (лежащая кпереди от турецкого седла), не имеющая сегментации, развивается, по-видимому, как новообразование в связи с увеличением размеров переднего мозга.
Филогенетически мозговой череп прошел три стадии развития: перепончатую, хрящевую и костную.
У круглоротых он практически весь перепончатый и не имеет передней, несегментированной, части.
Череп хрящевых рыб почти полностью хрящевой, причем включает в себя как заднюю, первично сегментированную, часть, так и переднюю.
У костных рыб и остальных позвоночных осевой череп становится костным за счет процессов окостенения хряща в области его основания (основная, клиновидная, решетчатая кости) и за счет возникновения покровных костей в верхней его части (теменные, лобные, носовые кости). Кости осевого черепа в процессе прогрессивной эволюции претерпевают олигомеризацию. Появление большого количества зон окостенения и последующее слияние их вместе при формировании таких костей, как лобная, височная и др., свидетельствуют об этом. Широко известны у человека такие аномалии мозгового черепа, как наличие межтеменных, а также двух лобных костей с метопическим швом между ними (рис. 14.8). Никакими патологическими явлениями они не сопровождаются и обнаруживаются поэтому обычно случайно после смерти.
Висцеральный череп впервые появляется также у низших позвоночных. Он формируется из мезенхимы эктодермального происхождения, которая группируется в виде сгущений, имеющих форму дужек, в промежутках между жаберными щелями глотки. Первые две дужки получают особенно сильное развитие и дают начало челюстной и подъязычной дугам взрослых животных. Следующие дуги в числе 4-5 пар выполняют опорную функцию для жабр и называются жаберными.
У хрящевых рыб впереди челюстной дуги располагаются обычно еще 1-2 пары предчелюстных дуг, имеющих рудиментарный характер. Это свидетельствует о том, что у предков позвоночных имелось большее количество висцеральных дуг, чем б или 7, а их дифференцировка происходила на фоне олигомеризации.
Челюстная дуга состоит из двух хрящей. Верхний называют нёбно-квадратным, он выполняет функцию первичной верхней челюсти. Нижний, или меккелев, хрящ - первичная нижняя челюсть. На вентральной стороне глотки меккелевы хрящи соединены друг с другом таким образом, что челюстная дуга кольцом охватывает ротовую полость. Вторая висцеральная дуга с каждой стороны состоит из гиомандибулярного хряща, сращенного с основанием мозгового черепа, и гиоида, соединенного с меккелевым хрящом. Таким образом, у хрящевых рыб обе первичные челюсти соединены с осевым черепом через вторую висцеральную дугу, в которой гиомандибулярный хрящ выполняет роль подвеска к мозговому черепу. Такой тип соединения челюстей и осевого черепа называют гиостильным (рис. 14.9).
У костных рыб начинается замещение первичных челюстей вторичными, состоящими из накладных костей - челюстной и предчелюстной сверху и зубной внизу. Нёбно-квадратный и меккелев хрящи при этом уменьшаются в размерах и смещаются кзади. Гиомандибулярный хрящ продолжает выполнять функции подвеска, поэтому череп остается гиостильным.
Земноводные в связи с переходом к наземному существованию претерпели значительные изменения висцерального черепа. Жаберные дуги частично редуцируются, а частично, меняя функции, входят в состав хрящевого аппарата гортани. Челюстная дуга своим верхним элементом - нёбно-квадратным хрящом - срастается полностью с основанием мозгового черепа, и череп становится, таким образом, аутостильным. Гиомандибулярный хрящ, сильно редуцированный и освободившийся от функции подвеска, располагаясь в области первой жаберной щели внутри слуховой капсулы, взял на себя функцию слуховой косточки - столбика,- передающей звуковые колебания от наружного к внутреннему уху.
Висцеральный череп пресмыкающихся также аутостилен. Для челюстного аппарата характерна более высокая степень окостенения, чем у земноводных. Часть хрящевого материала жаберных дуг входит в состав не только гортани, но и трахеи.
Нижняя челюсть млекопитающих сочленяется с височной костью сложным суставом, позволяющим не только захватывать пищу, но и совершать сложные жевательные движения.
Одна слуховая косточка - столбик,- характерная для земноводных и пресмыкающихся, уменьшаясь в размерах, превращается в стремечко, а рудименты нёбно-квадратного и меккелева хрящей, полностью выходящие из состава челюстного аппарата, преобразуются соответственно в наковаленку и молоточек. Таким образом, создается единая функциональная цепь из трех слуховых косточек в среднем ухе, характерная только для млекопитающих (рис. 14.9).

Рис. 14.9. Эволюция двух первых висцеральных жаберных дуг позвоночных.
А- хрящевая рыба; Б- земноводное; В- пресмыкающееся; Г- млекопитающее:
1 -нёбно-квадратный хрящ, 2-меккелев хрящ, 3- гиомандибулярный хрящ, 4-гиоид, 5- столбик, 6- накладные кости вторичных челюстей, 7-наковаленка, 8- стремечко, 9- молоточек; гомологичные образования обозначены соответствующей штриховкой
Рекапитуляция основных этапов филогенеза висцерального черепа происходит и в онтогенезе человека. Нарушение дифференцировки элементов челюстной жаберной дуги в слуховые косточки является механизмом формирования такого порока развития среднего уха, как расположение в барабанной полости только одной слуховой косточки - столбика, что соответствует строению звукопередающего аппарата земноводных и пресмыкающихся.
Скелет конечностей
У хордовых выделяются непарные и парные конечности. Непарные (спинные, хвостовой" и анальный плавники) являются основными органами передвижения у бесчерепных, рыб и в меньшей степени у хвостатых амфибий. У рыб возникают также парные конечности - грудные и брюшные плавники, на базе которых впоследствии развиваются парные конечности наземных четвероногих животных.
Подробнее рассмотрим происхождение и эволюцию парных конечностей.
У личинок рыб, а также у современных бесчерепных вдоль тела с обеих сторон тянутся боковые кожные складки, называемые метаплевральными (рис. 14.10). Они не имеют ни скелета, ни собственной мускулатуры, выполняя пассивную роль - стабилизацию положения тела и увеличение площади брюшной поверхности, облегчающие перемещение в водной среде. Вероятно, у предков рыб, переходящих к более активному образу жизни, в этих складках появились мышечные элементы и хрящевые лучи, связанные с сомитами по происхождению и поэтому расположенные метамерно. Такие складки, приобретя подвижность, могут выполнять роль рулей глубины, однако для изменения положения тела в пространстве большее значение имеют их передние и задние отделы, как наиболее удаленные от центра тяжести. Поэтому эволюция шла по пути интенсификации функций крайних отделов и ослабления функций центральных частей.

Рис. 14.10. Формирование передних и задних конечностей из метаплевральных складок: I -III -гипотетические этапы эволюции
В результате из передних отделов складок развились грудные, а из задних - брюшные плавники (рис. 14.10). Не исключено, что формированию только двух пар конечностей на боковых сторонах тела предшествовал распад сплошных складок на ряд парных плавников, большее значение из которых также имели передние и задние. Об этом свидетельствует существование ископаемых остатков древнейших низкоорганизованных рыб с многочисленными плавниками (рис. 14.11). За счет слияния оснований хрящевых лучей возникли плечевой и тазовый пояса. Остальные их участки дифференцировались в скелет свободных конечностей.

Рис. 14.11. Древняя акулообразная рыба с многочисленными парными конечностями
У большинства рыб в скелете парных плавников выделяют проксимальный отдел, состоящий из небольшого числа хрящевых или костных пластинок, и дистальный, в состав которого входит большое количество расположенных радиально сегментированных лучей. С поясами конечностей плавники соединены малоподвижно. Они не могут служить опорой телу при передвижении по дну или суше. У кистеперых рыб скелет парных конечностей имеет иное строение. Общее количество их костных элементов уменьшено, и они имеют более крупные размеры. Проксимальный отдел состоит только из одного крупного костного элемента, соответствующего плечевой или бедренной костям передних или задних конечностей. Далее следуют две более мелкие косточки, гомологичные локтевой и лучевой или большой и малой берцовым костям. На них опираются 7-12 радиально расположенных лучей. В соединении с поясами конечностей у такого плавника участвуют только гомологи плечевой или бедренной костей, поэтому плавники кистеперых рыб оказываются активно подвижными (рис. 14.12, А, Б ) и могут использоваться не только для изменения направления движения в воде, но и для перемещения по твердому субстрату.
Жизнь этих рыб в мелких пересыхающих водоемах в девонском периоде способствовала отбору форм с более развитыми и подвижными конечностями. Наличие у них добавочных органов дыхания (см. разд. 14.3.4) стало второй предпосылкой выхода на сушу и возникновения других адаптации к наземному существованию, результатом чего явилось происхождение земноводных и всей группы Tetrapoda. Первые их представители - стегоцефалы - обладали семи- и пятипалыми конечностями, сохраняющими сходство с плавниками кистеперых рыб (рис. 14.12, B )

Рис. 14.12. Скелет конечности кистеперой рыбы (А ), его основание (Б ) и скелет передней лапы стегоцефала (В ): I- плечевая кость, 2-локтевая кость, 3- лучевая кость
В скелете запястья сохранено правильное радиальное расположение костных элементов в 3-4 ряда, в пястье располагается 7-5 костей, а далее также радиально лежат фаланги 7-5 пальцев.
У современных земноводных количество пальцев в конечностях равно пяти или происходит их олигомеризация до четырех.
Дальнейшее прогрессивное преобразование конечностей выражается в увеличении степени подвижности соединений костей, в уменьшении количества костей в запястье сначала до трех рядов у амфибий и затем до двух - у пресмыкающихся и млекопитающих. Параллельно уменьшается также и количество фаланг пальцев. Характерно также удлинение проксимальных отделов конечности и укорочение дистальных.
Расположение конечностей в ходе эволюции также меняется. Если у рыб грудные плавники находятся на уровне первого позвонка и обращены в стороны, то у наземных позвоночных в результате усложнения ориентации в пространстве появляется шея и возникает подвижность головы, а у пресмыкающихся и особенно у млекопитающих в связи с приподнятием тела над землей передние конечности перемещаются кзади и ориентируются не горизонтально, а вертикально. То же касается и задних конечностей.
Многообразие условий обитания, предоставляемых наземным образом жизни, обеспечивает многообразие форм передвижения: прыжки, бег, ползание, полет, рытье, лазание по скалам и деревьям, а при возвращении в водную среду - и плавание. Поэтому у наземных позвоночных можно встретить как почти неограниченное многообразие конечностей, так и их полную вторичную редукцию, причем многие сходные адаптации конечностей в разнообразных средах многократно возникали конвергентно (рис. 14.13). Однако в процессе онтогенеза у большинства наземных позвоночных проявляются общие черты в развитии конечностей: закладка их зачатков в виде малодифференцированных складок, формирование в кисти и стопе вначале шести или семи зачатков пальцев, крайние из которых вскоре редуцируются и в дальнейшем развивается только пять (рис. 14.14).

Рис. 14.13. Скелет передней конечности наземных позвоночных. А -лягушка- Б -саламандра; В -крокодил; Г -летучая мышь; Д -человек: 1 -плечевая кость, 2-лучевая кость, 3 -кости запястья, 4 -пясти, 5 -фаланги пальцев, 6 -локтевая кость
Рис. 14.14. Строение развивающейся конечности позвоночного: рр - prepollex, pin - postminimus - дополнительные рудиментарные I и VII пальцы
Интересно, что в эмбриогенезе высших позвоночных рекапитулирует не только строение конечностей предков, но и процесс их гетеротопии. Так, у человека верхние конечности закладываются на уровне 3-4-го шейных позвонков, а нижние - на уровне поясничных позвонков. В это же время конечности получают иннервацию из соответствующих отделов спинного мозга. Гетеротопия конечностей сопровождается формированием шейного, поясничного и крестцового нервных сплетений, нервы которых связаны с одной стороны с теми сегментами спинного мозга, из которых они вырастали в момент формирования конечностей, а с другой - с конечностями, переместившимися на новое место (рис. 14.15; см. также разд. 14.2.2.2).
В онтогенезе человека возможны многочисленные нарушения, ведущие к формированию врожденных пороков развития конечностей атавистического плана. Так, полидактилия, или увеличение количества пальцев, наследующаяся как аутосомно-доминантный признак, является результатом развития закладок дополнительных пальцев, характерных в норме для далеких предковых форм. Известен феномен полифалангии, характеризующийся увеличением числа фаланг обычно большого пальца кисти. В основе его возникновения - развитие трех фаланг в первом пальце, как это в норме наблюдается у пресмыкающихся и земноводных с недифференцированными пальцами конечностей. Двусторонняя полифалангия наследуется аутосомно-доминантно.
Серьезным пороком развития является нарушение гетеротопии пояса верхних конечностей из шейной области на уровень 1-2-го грудных позвонков. Эту аномалию называют болезнью Шпренгеля или врожденным высоким стоянием лопатки (рис. 14.16). Она выражается в том, что плечевой пояс с одной либо с двух сторон находится выше нормального положения на несколько сантиметров. В связи с тем что такое нарушение часто сопровождается аномалиями рёбер, грудного отдела позвоночника и деформацией лопаток, следует думать, что механизмы его возникновения - не только нарушение перемещения органов, но и обусловленное этим нарушение морфогенетических корреляций (см. § 13.4).
Сравнительно-анатомический обзор эволюции скелета хордовых свидетельствует о том, что скелет человека полностью гомологичен опорному аппарату предковых и родственных форм. Поэтому многие пороки его развития у человека можно объяснить родством млекопитающих с пресмыкающимися, земноводными и рыбами. Однако в процессе антропогенеза появились такие особенности скелета, которые характерны лишь для человека и связаны с его прямохождением и трудовой деятельностью. К ним относят: 1) изменения стопы переставшей выполнять хватательную функцию, выражающиеся в потере способности к противопоставлению большого пальца и появлению ее сводов, служащих для амортизации при ходьбе; 2) изменения позвоночного столба - его S-образный изгиб, обеспечивающий пластичность движений в вертикальном положении; 3) изменения черепа - резкое уменьшение его лицевой части и увеличение мозговой, смещение большого затылочного отверстия кпереди, увеличение сосцевидного отростка и сглаживание затылочного рельефа, к которому прикрепляются мышцы шеи и выйная связка; 4) специализация верхних (передних) конечностей как органа труда; 5) появление подбородочного выступа в связи с развитием членораздельной речи.

Рис. 14.15. Формирование передних конечностей, их гетеротопия и иннервация в онтогенезе человека. А -врастание шейных миотомов в формирующуюся переднею конечность зародыша; Б -развитие кожной иннервации руки; В - расположение шейного и плечевого сплетений, участвующих в иннервации руки:
1 -шейные миотомы, 2- грудные миотомы, 3 -поясничные миотомы; буквами С, Т, L обозначены шейные, грудные и поясничные сегменты

Рис. 14.16. Болезнь Шпренгеля (пояснение см. в тексте)
Несмотря на то что становление анатомо-морфологических особенностей скелета у человека, по-видимому, завершено, адаптации к прямохождению у него имеют, как и все адаптации вообще, относительный характер. Так, при большой физической нагрузке возможно смещение позвонков или межпозвонковых дисков. Человек, перейдя к прямохождению, утратил способность к быстрому бегу и передвигается значительно медленнее большинства четвероногих животных.
Естественно, что в ходе внутриутробного развития черты скелета, характеризующие человека как уникальный биологический вид, формируются на конечных его этапах либо даже, как, например, S-образная форма позвоночника, в раннем постнатальном периоде развития. Они являются фактически анаболиями, возникшими в ходе филогенеза приматов. Поэтому атавистические аномалии скелета, связанные с задержками развития признаков, характерных только для человека, встречаются наиболее часто. Они практически не снижают жизнеспособность, но дети, обладающие ими, нуждаются в ортопедической коррекции, гимнастике и массаже. К таким аномалиям относят легкие формы врожденного плоскостопия, косолапости, узкую грудную клетку, отсутствие подбородочного выступа и некоторые другие.
Мышечная система
У представителей типа Хордовые мускулатура подразделяется по характеру развития и иннервации на соматическую и висцеральную.
Соматическая мускулатура развивается из миотомов и иннервируется нервами, волокна которых выходят из спинного мозга в составе брюшных корешков спинномозговых нервов. Висцеральная мускулатура развивается из других участков мезодермы и иннервируется нервами вегетативной нервной системы. Вся соматическая мускулатура поперечно-полосата, а висцеральная может быть как поперечно-полосатой, так и гладкой (рис. 14.17).

Рис. 14.17. Соматическая и висцеральная мускулатура позвоночных:
1 -соматическая мускулатура, развивающаяся из миотомов, 2- висцеральная мускулатура жаберной области
Висцеральная мускулатура
Наиболее существенные изменения претерпела висцеральная мускулатура, связанная с висцеральными дугами передней части пищеварительной трубки. У низших позвоночных большая часть этой мускулатуры представлена общим сжимателем висцерального аппарата - m. constrictor superficialis, покрывающим всю область жаберных дуг со всех сторон. В области челюстной дуги эта мышца иннервируется тройничным нервом (V), в области подъязычной дуги - лицевым (VII), в области первой жаберной дуги - языкоглоточным (IX), наконец, ее часть, лежащая каудальнее,- блуждающим нервом (X). В связи с этим все производные соответствующих висцеральных дуг и мышц, связанных с ними, иннервируются впоследствии у всех позвоночных перечисленными нервами.
В передней части сжимателя обособляется крупная мускулатурная масса, обслуживающая челюстной аппарат. Позади висцерального аппарата дифференцируется трапециевидная мышца m. trapezius, отдельными пучками прикрепляющаяся к последним жаберным щелям и переднему краю спинного отдела плечевого пояса. Часть поверхностного сжимателя в области подъязычной дуги у пресмыкающихся разрастается, охватывает шею снизу и с боков и образует сжиматель шеи m. sphincter colli. У млекопитающих эта мышца делится на два слоя: глубокий и поверхностный. Глубокий сохраняет прежнее название, а поверхностный называется platysma myoides и располагается подкожно. Эти две мышцы разрастаются на всю область головы и дают начало сложной системе лицевой подкожной мускулатуры, которую у приматов и человека называют мимической. Поэтому вся мимическ